

Plant Defense Responses to Insect Herbivores Through Molecular Signaling, Secondary Metabolites, and Associated Epigenetic Regulation
- PMID: 39959634
- PMCID: PMC11830398
- DOI: 10.1002/pei3.70035
Plant Defense Responses to Insect Herbivores Through Molecular Signaling, Secondary Metabolites, and Associated Epigenetic Regulation
Abstract
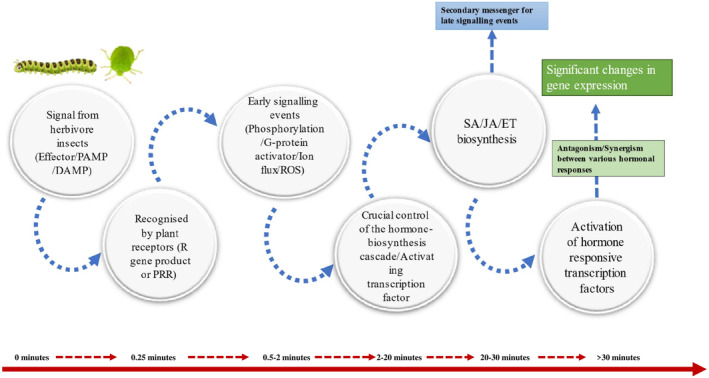
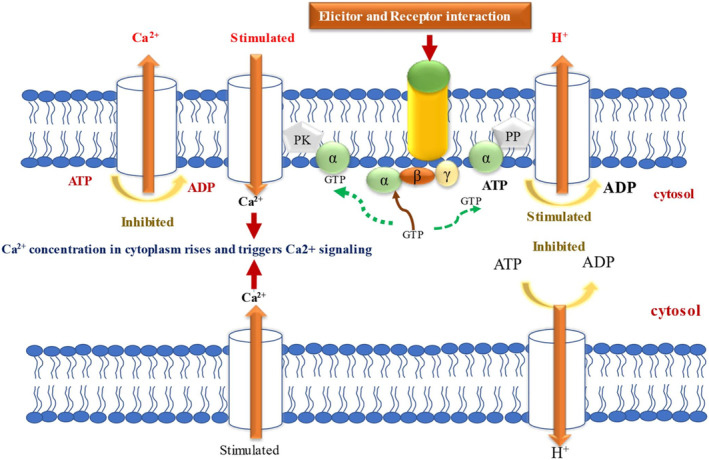
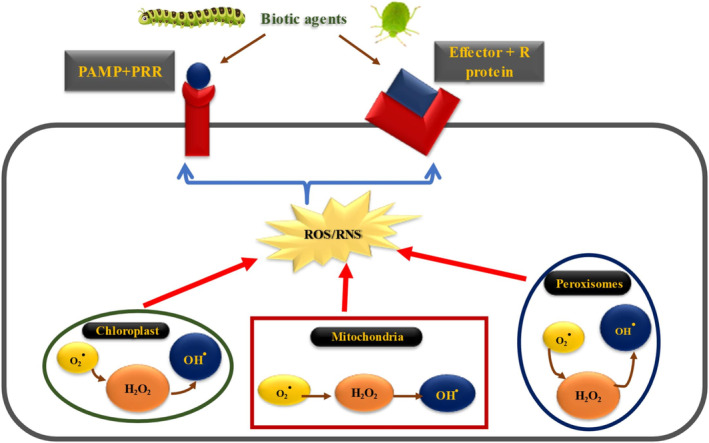
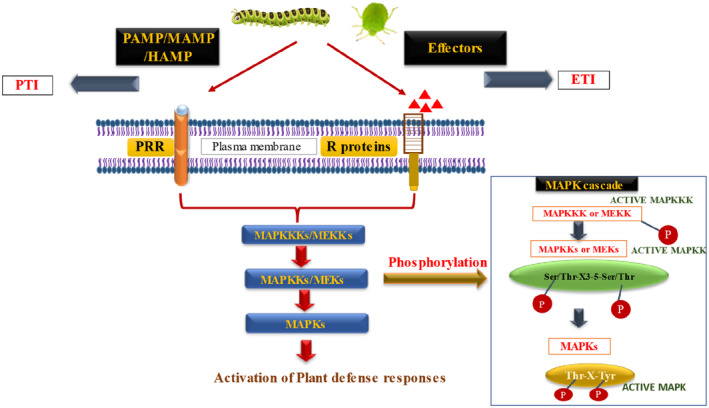
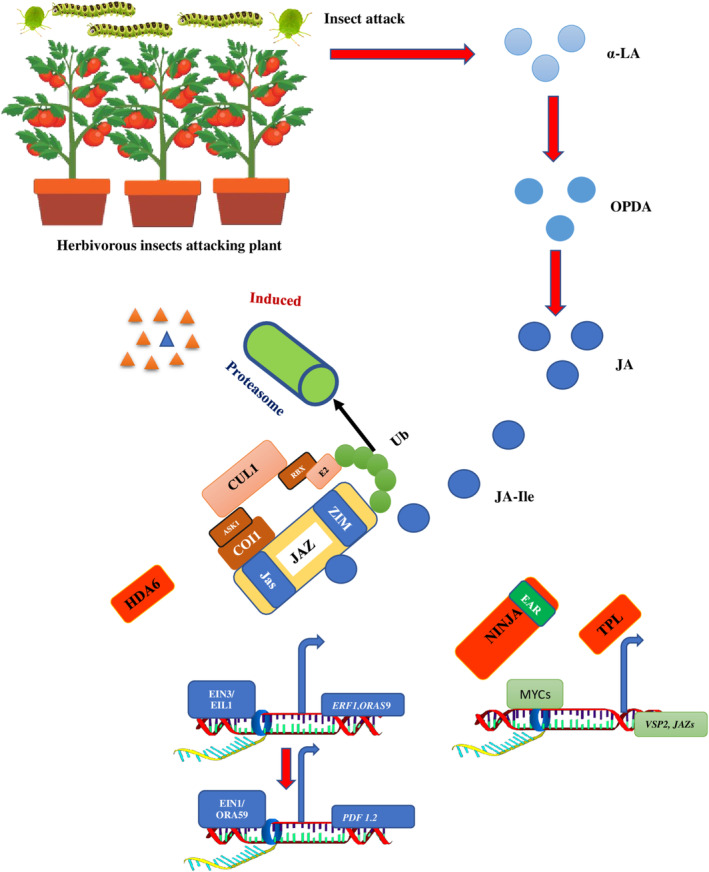
Over millions of years of interactions, plants have developed complex defense mechanisms to counteract diverse insect herbivory strategies. These defenses encompass morphological, biochemical, and molecular adaptations that mitigate the impacts of herbivore attacks. Physical barriers, such as spines, trichomes, and cuticle layers, deter herbivores, while biochemical defenses include the production of secondary metabolites and volatile organic compounds (VOCs). The initial step in the plant's defense involves sensing mechanical damage and chemical cues, including herbivore oral secretions and herbivore-induced VOCs. This triggers changes in plasma membrane potential driven by ion fluxes across plant cell membranes, activating complex signal transduction pathways. Key hormonal mediators, such as jasmonic acid, salicylic acid, and ethylene, orchestrate downstream defense responses, including VOC release and secondary metabolites biosynthesis. This review provides a comprehensive analysis of plant responses to herbivory, emphasizing early and late defense mechanisms, encompassing physical barriers, signal transduction cascades, secondary metabolites synthesis, phytohormone signaling, and epigenetic regulation.
Keywords: epigenetic regulations; herbivore attack; molecular signaling; plant–herbivore interaction; secondary metabolites.
© 2025 The Author(s). Plant‐Environment Interactions published by New Phytologist Foundation and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Adithya, A. , Indu Rani C., Savitha B. K., Murugan M., Sudha M., and Prabhu M.. 2024. “Harnessing the Crop Wild Relatives in Genetic Improvement of Eggplant, Potato, and Tomato.” Genetic Resources and Crop Evolution: 1–17. 10.1007/s10722-024-01565-8. - DOI
-
- Afentoulis, D. G. , Cusumano A., Greenberg L. O., Caarls L., and Fatouros N. E.. 2021. “Attraction of Trichogramma Wasps to Butterfly Oviposition‐Induced Plant Volatiles Depends on Brassica Species, Wasp Strain and Leaf Necrosis.” Frontiers in Ecology and Evolution 9: 703134. 10.3389/fevo.2021.703134. - DOI
-
- Ahmad, M. A. , Kumar P., Mahanta D. K., and Rai C. P.. 2023. “Physical and Biochemical Parameters Associated With Resistance to Callosobruchus chinensis Linn. In Chickpea.” Pharma Innovation Journal 12, no. 6: 728–733.
Publication types
LinkOut - more resources
Full Text Sources