

Small molecule-mediated inhibition of the oxidoreductase ERO1A restrains aggressive breast cancer by impairing VEGF and PD-L1 in the tumor microenvironment
- PMID: 39962052
- PMCID: PMC11833095
- DOI: 10.1038/s41419-025-07426-1
Small molecule-mediated inhibition of the oxidoreductase ERO1A restrains aggressive breast cancer by impairing VEGF and PD-L1 in the tumor microenvironment
Abstract
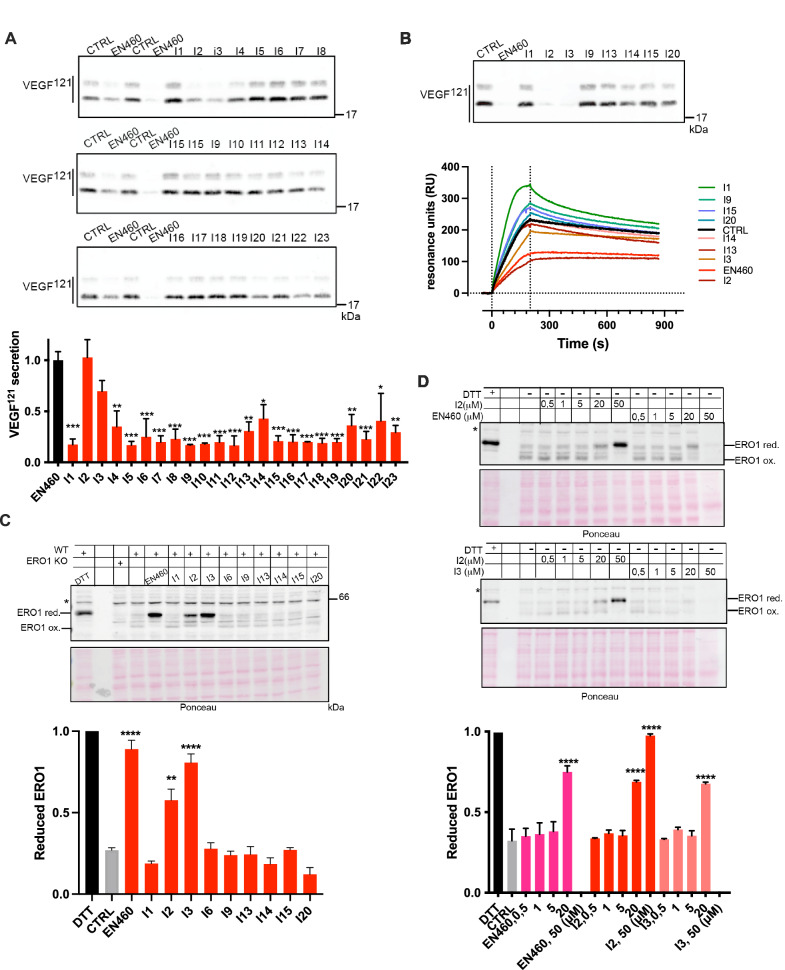
Cancer cells adapt to harsh environmental conditions by inducing the Unfolded Protein Response (UPR), of which ERO1A is a mediator. ERO1A aids protein folding by acting as a protein disulfide oxidase, and under cancer-related hypoxia conditions, it favors the folding of angiogenic VEGFA, leading tumor cells to thrive and spread. The upregulation of ERO1A in cancer cells, oppositely to the dispensability of ERO1A activity in healthy cells, renders ERO1A a perfect target for cancer therapy. Here, we report the upregulation of ERO1A in a cohort of aggressive triple-negative breast cancer (TNBC) patients in which ERO1A levels correlate with a higher risk of breast tumor recurrence and metastatic spread. For ERO1A target validation and therapy in TNBC, we designed new ERO1A inhibitors in a structure-activity campaign of the prototype EN460. Cell-based screenings showed that the presence of the Micheal acceptor in the compound is necessary to engage the cysteine 397 of ERO1A but not sufficient to set out the inhibitory effect on ERO1A. Indeed, the ERO1 inhibitor must adopt a non-coplanar rearrangement within the ERO1A binding site. I2 and I3, two new EN460 analogs with different phenyl-substituted moieties, efficiently inhibited ERO1A, blunting VEGFA secretion. Accordingly, in vitro assays to measure ERO1A engagement and inhibition confirmed that I2 and I3 bind ERO1A and restrain its activity with a IC50 in a low micromolar range. EN460, I2 and I3 triggered breast cancer cytotoxicity while specifically inhibiting ERO1A in a dose-dependent manner. I2 more efficiently impaired cancer-relevant features such as VEGFA secretion and related cell migration. I2 also acted on the tumor microenvironment and viability of xenografts and syngeneic TNBC. Thus, small molecule-mediated ERO1A pharmacological inhibition is feasible and promises to lead to effective therapy for the still incurable TNBC.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests. Ethics approval and consent to participate: Human studies were approved by the local ethical committee of “Azienda Socio-Sanitaria Territoriale of Cremona” (RaLCTrVs1Ott_2000). All patients participated in this study provided written informed consents in accordance with the Declaration of Helsinki. Mouse studies are reported in compliance with the ARRIVE guidelines and conducted in conformity with the following laws, regulations and policies governing the care and use of laboratory animals: Italian Governing Law (D.lgs 26/2014, authorization 19/2008-A issued 6 March 2008 by Ministry of Health; authorization 896/2023-PR to E. Zito); Mario Negri Institutional Regulations and Policies providing internal authorization for people conducting animal experiments (Quality Management System Certificate—UNI EN ISO9001: 2008—registration number 6121); EU directives and guidelines (EEC Council Directive 2010/63/UE). Consent for publication: Not applicable.
Figures







References
-
- Manuelli V, Pecorari C, Filomeni G, Zito E. Regulation of redox signaling in HIF-1-dependent tumor angiogenesis. FEBS J. 2021. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
