

Mitochondrial defects and metabolic vulnerabilities in Lynch syndrome-associated MSH2-deficient endometrial cancer
- PMID: 39964762
- PMCID: PMC11949016
- DOI: 10.1172/jci.insight.185946
Mitochondrial defects and metabolic vulnerabilities in Lynch syndrome-associated MSH2-deficient endometrial cancer
Abstract
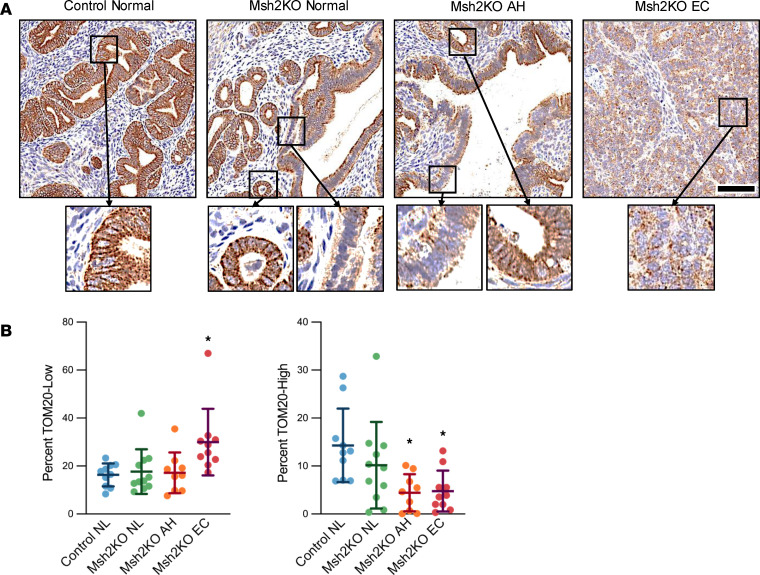
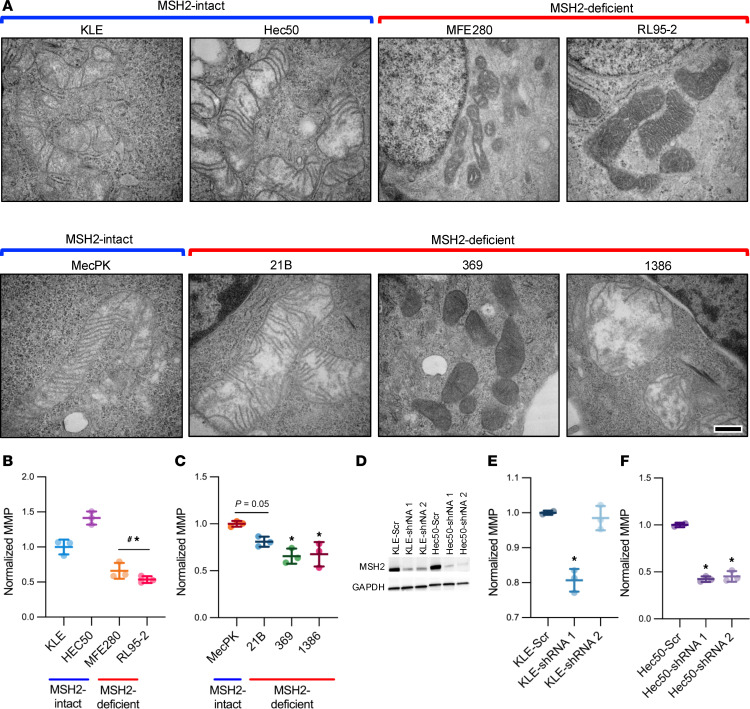
Lynch syndrome (LS), caused by inherited mutations in DNA mismatch repair genes, including MSH2, carries a 60% lifetime risk of developing endometrial cancer (EC). Beyond hypermutability, mechanisms driving LS-associated EC (LS-EC) remain unclear. We investigated MSH2 loss in EC pathogenesis using a mouse model (PR-Cre Msh2LoxP/LoxP, abbreviated Msh2KO), primary cell lines, human tissues, and human EC cells with isogenic MSH2 knockdown. By 8 months, 58% of Msh2KO mice developed endometrial atypical hyperplasia (AH), a precancerous lesion. At 12-16 months, 50% of Msh2KO mice exhibited either AH or ECs with histologic similarities to human LS-ECs. Transcriptomic profiling of EC from Msh2KO mice revealed mitochondrial dysfunction-related pathway changes. Subsequent studies in vitro and in vivo revealed mitochondrial dysfunction based on 2 mechanisms: mitochondrial content reduction and structural disruptions in retained mitochondria. Human LS-ECs also exhibited mitochondrial content reduction compared with non-LS-ECs. Functional studies demonstrated metabolic reprogramming of MSH2-deficient EC, including reduced oxidative phosphorylation and increased susceptibility to glycolysis suppression. These findings identified mitochondrial dysfunction and metabolic disruption as consequences of MSH2 deficiency in EC. Mitochondrial and metabolic aberrations should be evaluated as biomarkers for endometrial carcinogenesis or risk stratification and represent potential targets for cancer interception in women with LS.
Keywords: Cancer; Cell biology; Mitochondria; Obstetrics/gynecology; Oncology.
Conflict of interest statement
Figures





Update of
-
Mitochondrial defects and metabolic vulnerabilities in Lynch syndrome-associated MSH2-deficient endometrial cancer.bioRxiv [Preprint]. 2024 Jun 11:2024.06.10.596841. doi: 10.1101/2024.06.10.596841. bioRxiv. 2024. Update in: JCI Insight. 2025 Feb 18;10(6):e185946. doi: 10.1172/jci.insight.185946. PMID: 38915709 Free PMC article. Updated. Preprint.
References
-
- Idos G, Valle L, eds. Lynch Syndrome. University of Washington; 1993.
-
- Gruber SB, Thompson WD. A population-based study of endometrial cancer and familial risk in younger women. Cancer and Steroid Hormone Study Group. Cancer Epidemiol Biomarkers Prev. 1996;5(6):411–417. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
