

Amniotic epithelial Cell microvesicles uptake inhibits PBMCs and Jurkat cells activation by inducing mitochondria-dependent apoptosis
- PMID: 39967871
- PMCID: PMC11834128
- DOI: 10.1016/j.isci.2025.111830
Amniotic epithelial Cell microvesicles uptake inhibits PBMCs and Jurkat cells activation by inducing mitochondria-dependent apoptosis
Abstract
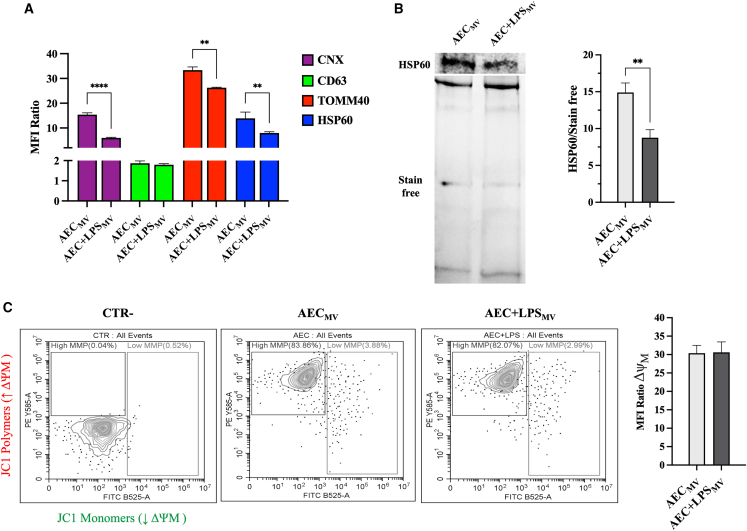
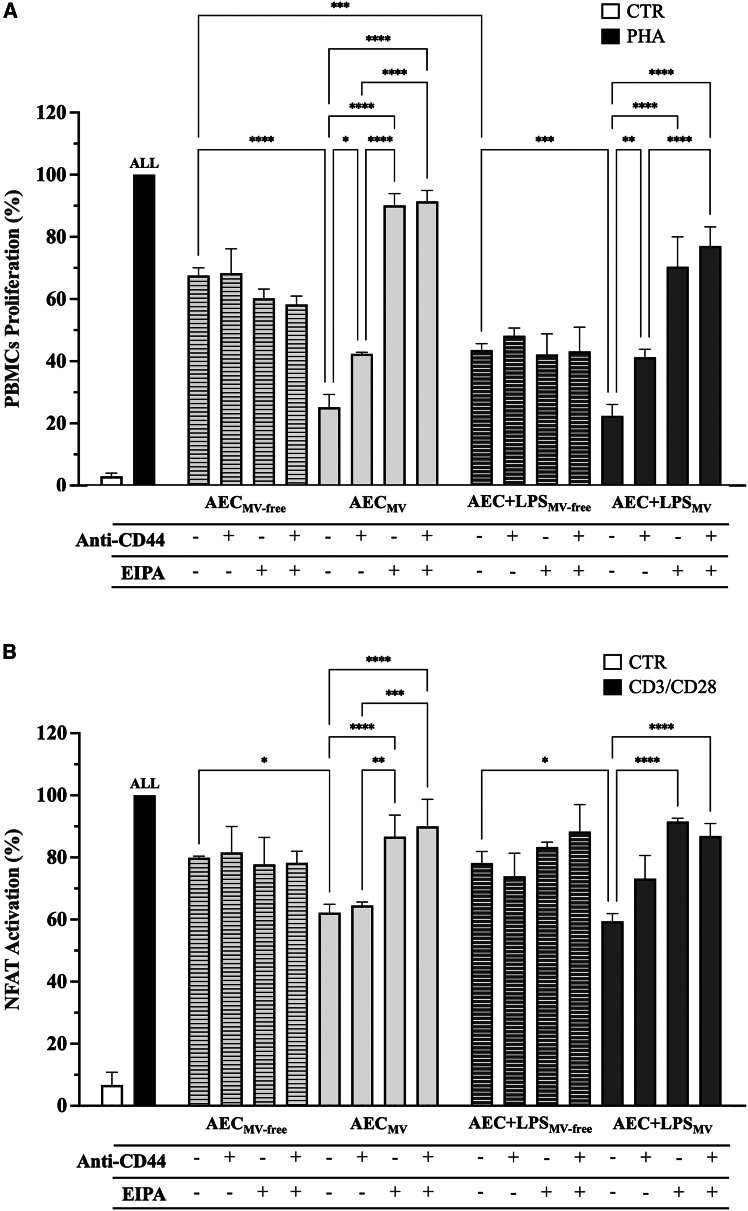
Amniotic epithelial cells (AECs) exhibit significant immunomodulatory and pro-regenerative properties, largely due to their intrinsic paracrine functions that are currently harnessed through the collection of their secretomes. While there is increasing evidence of the role of bioactive components freely secreted or carried by exosomes, the bioactive cargo of AEC microvesicles (MVs) and their crosstalk with the immune cells remains to be fully explored. We showed that under intrinsic conditions or in response to LPS, AEC-derived MV carries components such as lipid-mediated signaling molecules, ER, and mitochondria. They foster the intra/interspecific mitochondrial transfer into immune cells (PBMCs and Jurkat cells) in vitro and in vivo on the zebrafish larvae model of injury. The internalization of MV cargoes through macropinocytosis induces hyperpolarization of PBMC mitochondrial membranes and triggers MV-mediated apoptosis. This powerful immune suppressive mechanism triggered by AEC-MV cargo delivery paves the way for controlled and targeted cell-free therapeutic approaches.
Keywords: Cell biology; Functional aspects of cell biology; Immunology.
© 2025 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures








References
-
- Di Nicola M., Carlo-Stella C., Magni M., Milanesi M., Longoni P.D., Matteucci P., Grisanti S., Gianni A.M. Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood. 2002;99:3838–3843. doi: 10.1182/blood.v99.10.3838. - DOI - PubMed
Associated data
LinkOut - more resources
Full Text Sources
