

Cdc42EP3-bound septin scaffolds promote actin polymerization
- PMID: 39971161
- PMCID: PMC11952830
- DOI: 10.1016/j.jbc.2025.108325
Cdc42EP3-bound septin scaffolds promote actin polymerization
Abstract
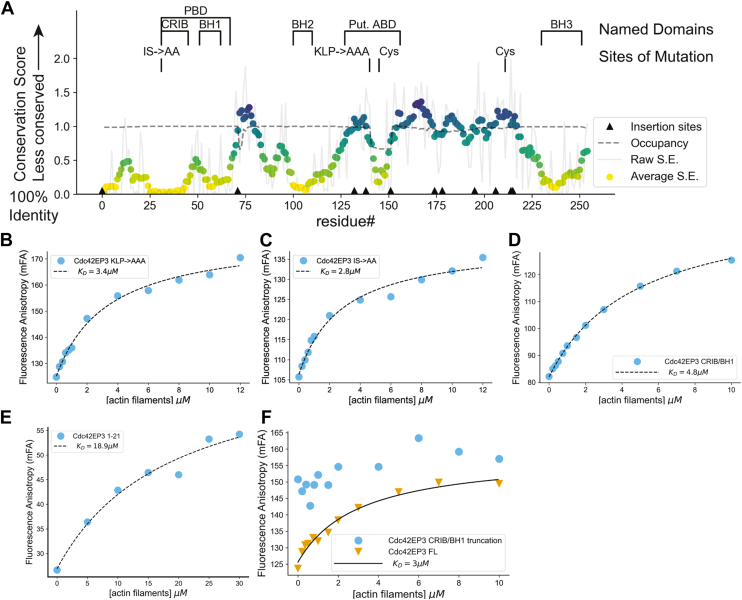
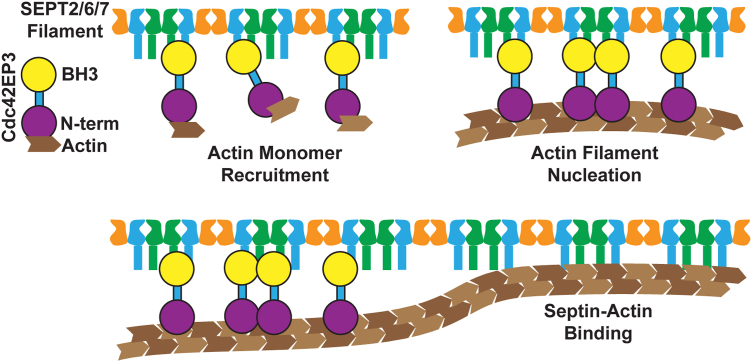
Septins are cytoskeletal filament-forming proteins that typically associate with membranes and perform critical functions in a variety of cellular processes. Septins often colocalize with actin and microtubule structures, yet our understanding of all the ways that septins contribute mechanistically to actin- and microtubule-based functions is incomplete. The Cdc42 effector protein Cdc42EP3 (also known as BORG2) promotes septin localization to actin structures in vivo, but little else is known about how Cdc42EP3 influences the interactions of septins and F-actin. Here, using purified components, we show that Cdc42EP3 binds directly to septins, actin filaments, and actin monomers. Moreover, septin-bound Cdc42EP3 accelerates actin filament polymerization. Thus, Cdc42EP3 is not merely a factor that crosslinks septins and F-actin, but one that promotes the formation of actin polymers along septin scaffolds.
Keywords: CDC42; Cdc42EP/BORG; G-actin; actin; bundling; cytoskeleton; filament nucleation; microfilaments; septin.
Copyright © 2025 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures





References
-
- Hartwell L.H. Genetic control of the cell division cycle in yeast. IV. Genes controlling bud emergence and cytokinesis. Exp. Cell Res. 1971;69:265–276. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
