

Human skeletal muscle fiber heterogeneity beyond myosin heavy chains
- PMID: 39971958
- PMCID: PMC11839989
- DOI: 10.1038/s41467-025-56896-6
Human skeletal muscle fiber heterogeneity beyond myosin heavy chains
Abstract
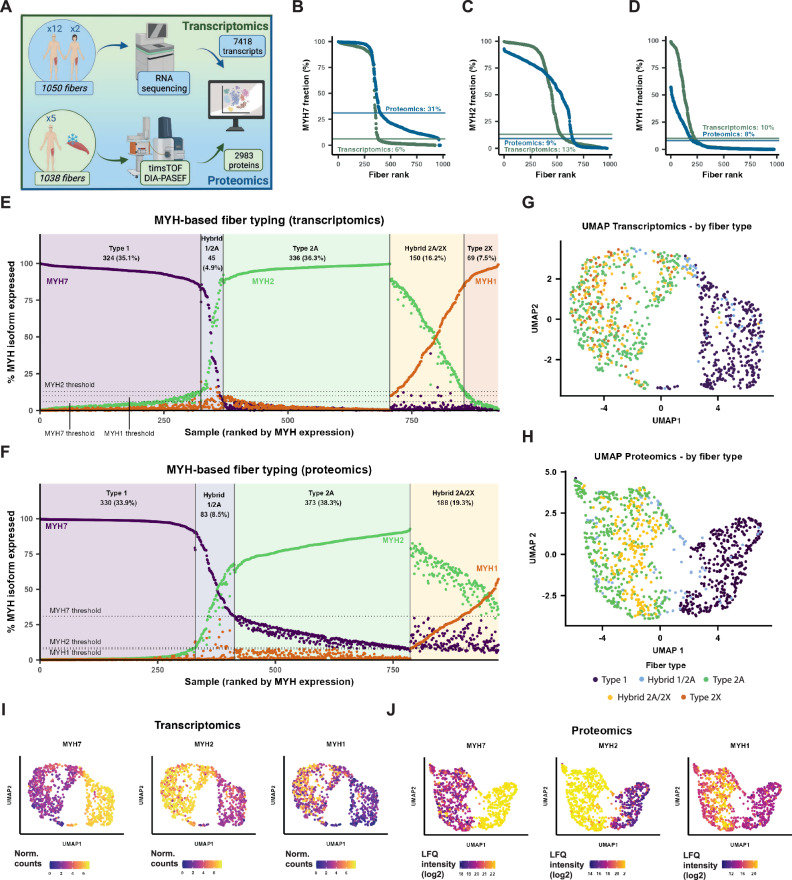
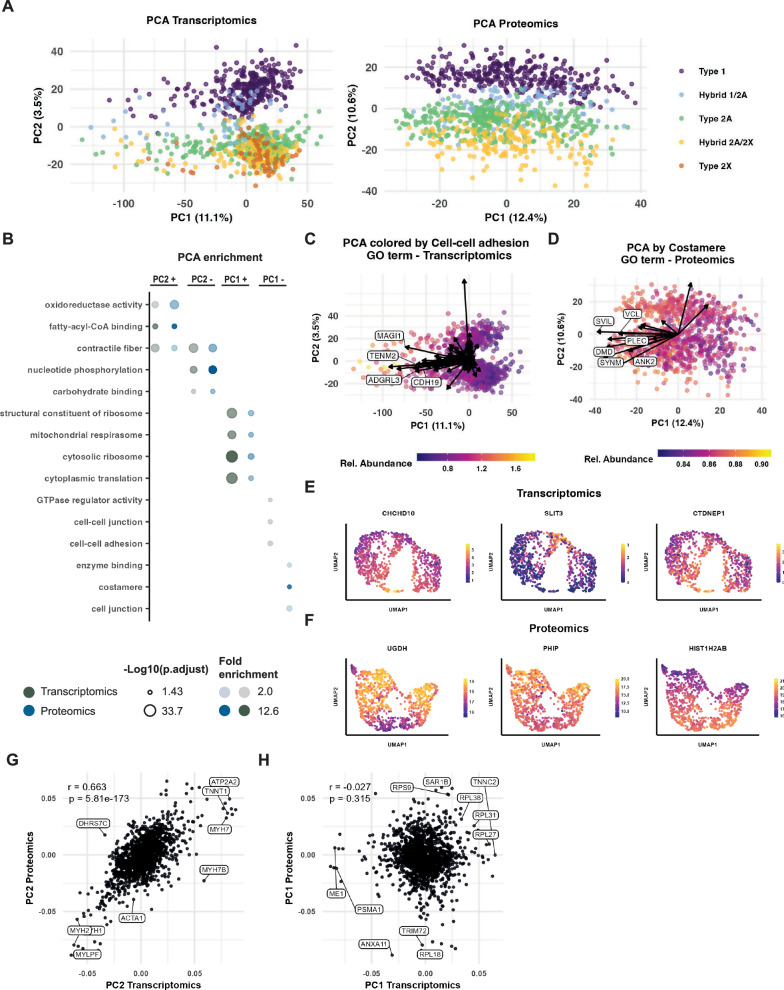
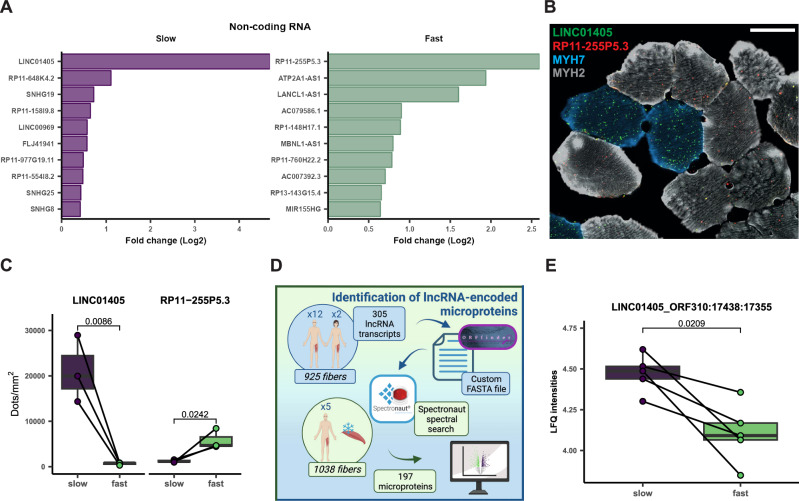
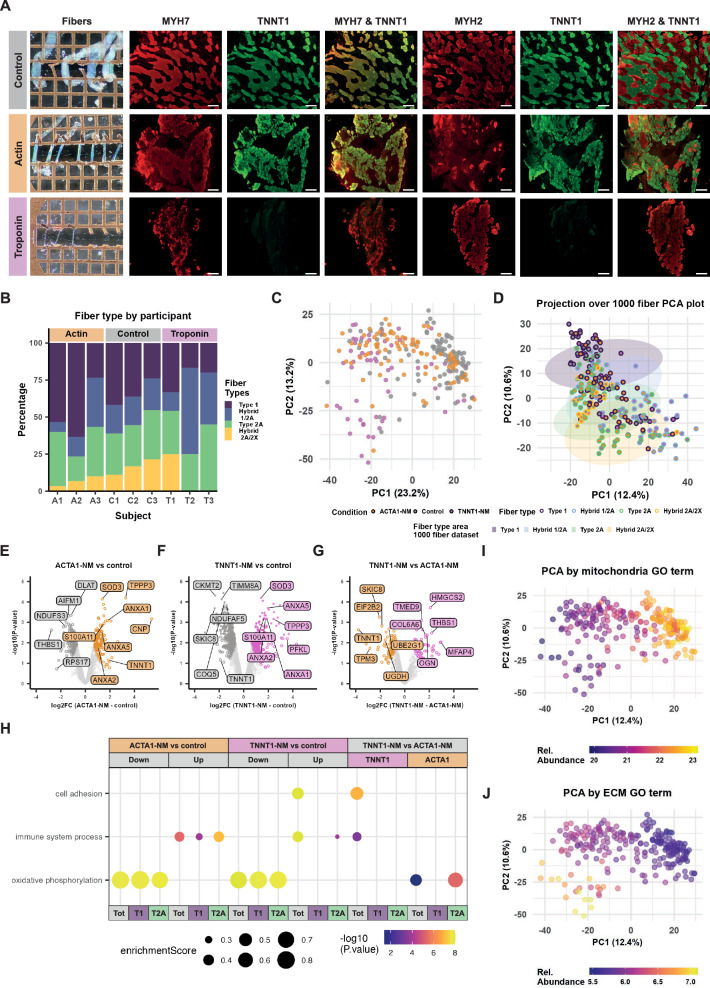
Skeletal muscle is a heterogenous tissue comprised primarily of myofibers, commonly classified into three fiber types in humans: one "slow" (type 1) and two "fast" (type 2A and type 2X). However, heterogeneity between and within traditional fiber types remains underexplored. We applied transcriptomic and proteomic workflows to 1050 and 1038 single myofibers from human vastus lateralis, respectively. Proteomics was conducted in males, while transcriptomics included ten males and two females. We identify metabolic, ribosomal, and cell junction proteins, in addition to myosin heavy chain isoforms, as sources of multi-dimensional variation between myofibers. Furthermore, whilst slow and fast fiber clusters are identified, our data suggests that type 2X fibers are not phenotypically distinct to other fast fibers. Moreover, myosin heavy chain-based classifications do not adequately describe the phenotype of myofibers in nemaline myopathy. Overall, our data indicates that myofiber heterogeneity is multi-dimensional with sources of variation beyond myosin heavy chain isoforms.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures


References
-
- Engel, W. K. The essentiality of histo- and cytochemical studies of skeletal muscle in the investigation of neuromuscular disease. 1962. Neurol.12, 778–784 (1962). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
