

GABAergic neuron-to-glioma synapses in diffuse midline gliomas
- PMID: 39972132
- PMCID: PMC11946904
- DOI: 10.1038/s41586-024-08579-3
GABAergic neuron-to-glioma synapses in diffuse midline gliomas
Abstract
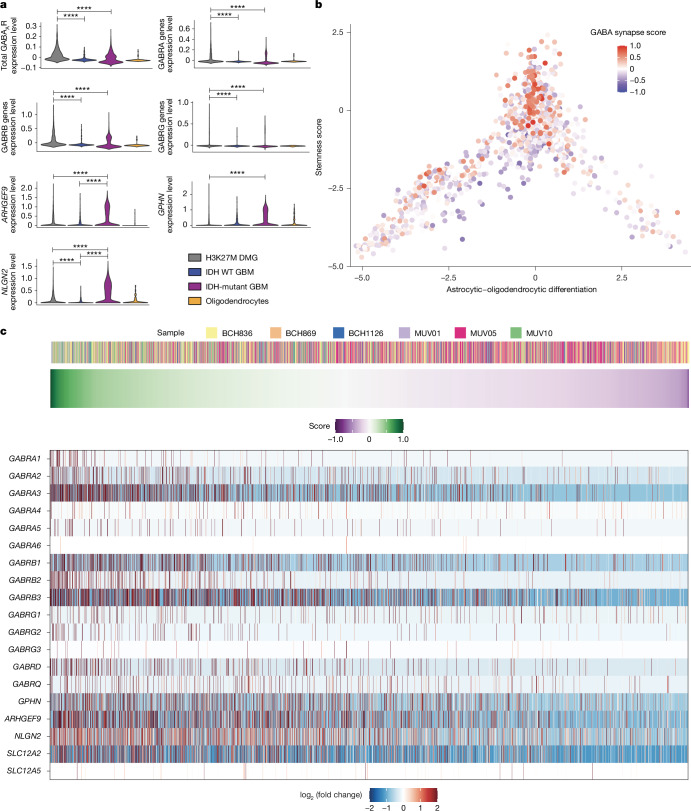
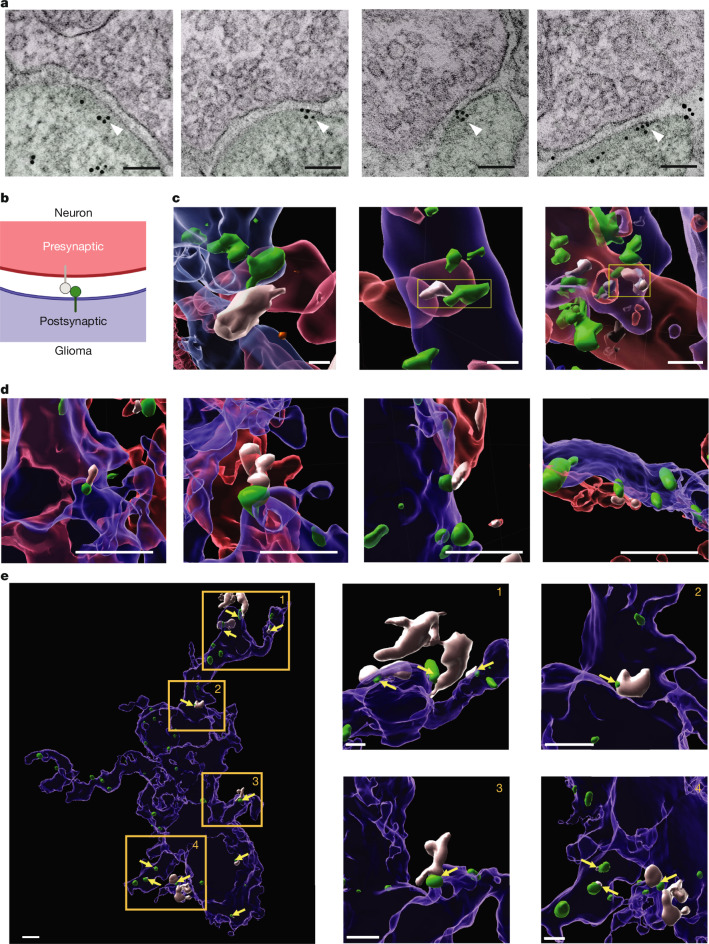
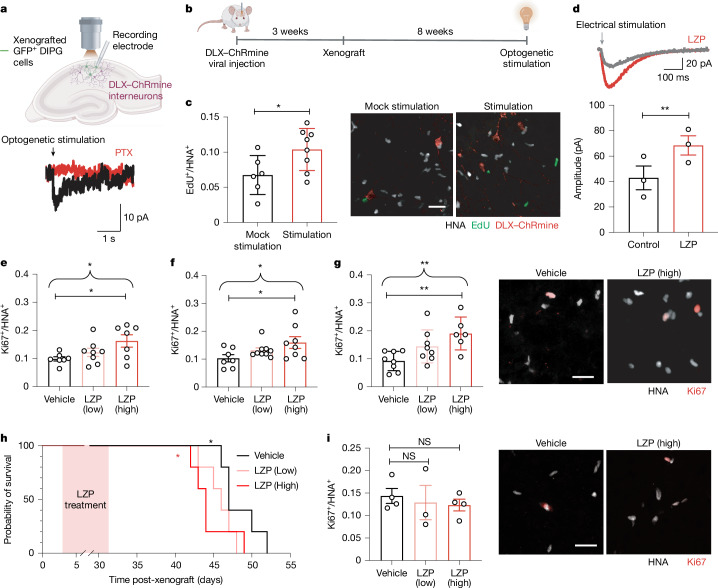
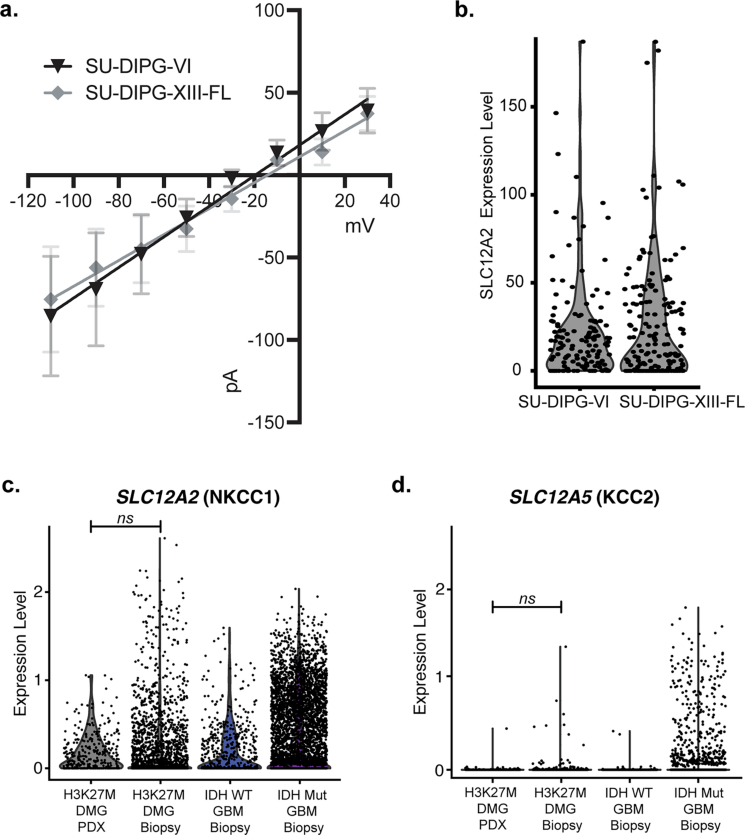
High-grade gliomas (HGGs) are the leading cause of brain cancer-related death. HGGs include clinically, anatomically and molecularly distinct subtypes that stratify into diffuse midline gliomas (DMGs), such as H3K27M-altered diffuse intrinsic pontine glioma, and hemispheric HGGs, such as IDH wild-type glioblastoma. Neuronal activity drives glioma progression through paracrine signalling1,2 and neuron-to-glioma synapses3-6. Glutamatergic AMPA receptor-dependent synapses between neurons and glioma cells have been demonstrated in paediatric3 and adult4 high-grade gliomas, and early work has suggested heterogeneous glioma GABAergic responses7. However, neuron-to-glioma synapses mediated by neurotransmitters other than glutamate remain understudied. Using whole-cell patch-clamp electrophysiology, in vivo optogenetics and patient-derived orthotopic xenograft models, we identified functional, tumour-promoting GABAergic neuron-to-glioma synapses mediated by GABAA receptors in DMGs. GABAergic input has a depolarizing effect on DMG cells due to NKCC1 chloride transporter function and consequently elevated intracellular chloride concentration in DMG malignant cells. As membrane depolarization increases glioma proliferation3,6, we found that the activity of GABAergic interneurons promotes DMG proliferation in vivo. The benzodiazepine lorazepam enhances GABA-mediated signalling, increases glioma proliferation and growth, and shortens survival in DMG patient-derived orthotopic xenograft models. By contrast, only minimal depolarizing GABAergic currents were found in hemispheric HGGs and lorazepam did not influence the growth rate of hemispheric glioblastoma xenografts. Together, these findings uncover growth-promoting GABAergic synaptic communication between GABAergic neurons and H3K27M-altered DMG cells, underscoring a tumour subtype-specific mechanism of brain cancer neurophysiology.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: M.M. holds equity in MapLight Therapeutics and Stellaromics, and stock in CARGO Therapeutics. The other authors declare no competing interests.
Figures







References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
