

This is a preprint.
Genes and Pathways Comprising the Human and Mouse ORFeomes Display Distinct Codon Bias Signatures that Can Regulate Protein Levels
- PMID: 39974974
- PMCID: PMC11838421
- DOI: 10.1101/2025.02.03.636209
Genes and Pathways Comprising the Human and Mouse ORFeomes Display Distinct Codon Bias Signatures that Can Regulate Protein Levels
Abstract
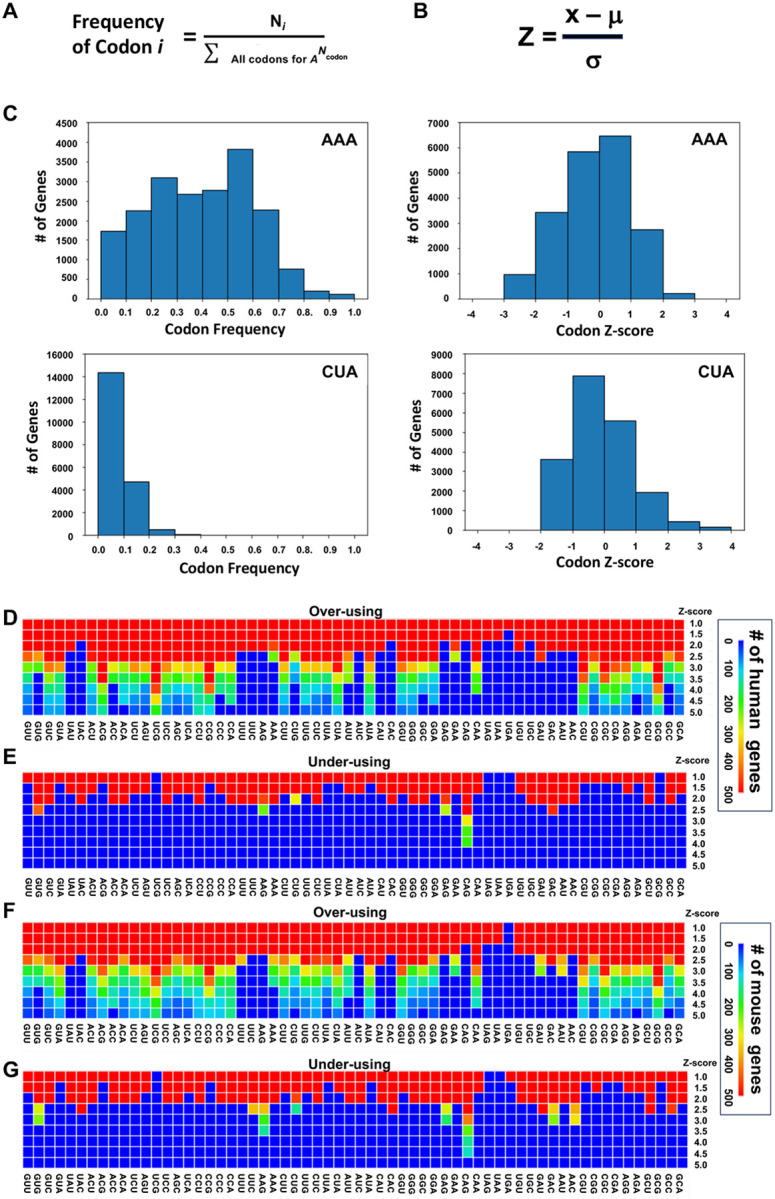
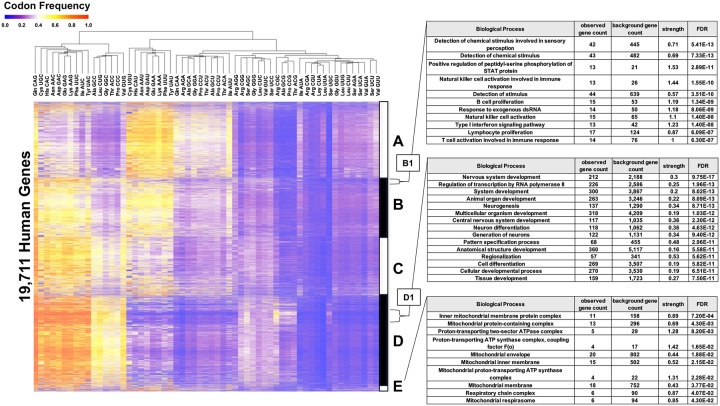
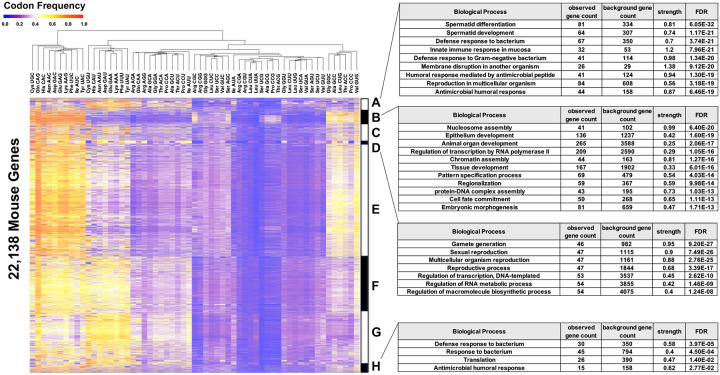
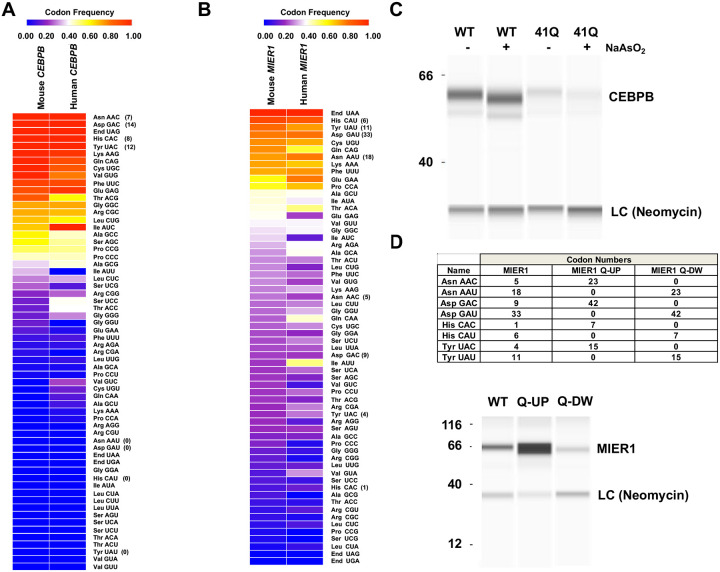
Arginine, glutamic acid and selenocysteine based codon bias has been shown to regulate the translation of specific mRNAs for proteins that participate in stress responses, cell cycle and transcriptional regulation. Defining codon-bias in gene networks has the potential to identify other pathways under translational control. Here we have used computational methods to analyze the ORFeome of all unique human (19,711) and mouse (22,138) open-reading frames (ORFs) to characterize codon-usage and codon-bias in genes and biological processes. We show that ORFeome-wide clustering of gene-specific codon frequency data can be used to identify ontology-enriched biological processes and gene networks, with developmental and immunological programs well represented for both humans and mice. We developed codon over-use ontology mapping and hierarchical clustering to identify multi-codon bias signatures in human and mouse genes linked to signaling, development, mitochondria and metabolism, among others. The most distinct multi-codon bias signatures were identified in human genes linked to skin development and RNA metabolism, and in mouse genes linked to olfactory transduction and ribosome, highlighting species-specific pathways potentially regulated by translation. Extreme codon bias was identified in genes that included transcription factors and histone variants. We show that re-engineering extreme usage of C- or U-ending codons for aspartic acid, asparagine, histidine and tyrosine in the transcription factors CEBPB and MIER1, respectively, significantly regulates protein levels. Our study highlights that multi-codon bias signatures can be linked to specific biological pathways and that extreme codon bias with regulatory potential exists in transcription factors for immune response and development.
Keywords: Codon bias; ORFeome; codon re-engineering; development; gene expression; queuosine; tRNA modification; transcription factors; translation.
Figures



References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources