

This is a preprint.
Chemotherapeutic 6-thio-2'-deoxyguanosine selectively targets and inhibits telomerase by inducing a non-productive telomere-bound telomerase complex
- PMID: 39975053
- PMCID: PMC11838547
- DOI: 10.1101/2025.02.05.636339
Chemotherapeutic 6-thio-2'-deoxyguanosine selectively targets and inhibits telomerase by inducing a non-productive telomere-bound telomerase complex
Abstract
Most cancers upregulate the telomere lengthening enzyme telomerase to achieve unlimited cell division. How chemotherapeutic nucleoside 6-thio-2'-deoxyguanosine (6-thio-dG) targets telomerase to inhibit telomere maintenance in cancer cells and tumors was unclear. Here, we demonstrate that telomerase insertion of 6-thio-dGTP prevents synthesis of additional telomeric repeats but does not disrupt telomerase binding to telomeres. Specifically, 6-thio-dG inhibits telomere extension after telomerase translocates along its product DNA to reposition the template, inducing a non-productive complex rather than enzyme dissociation. Furthermore, we provide direct evidence that 6-thio-dG treatment inhibits telomere synthesis by telomerase in cancer cells. In agreement, telomerase-expressing cancer cells harboring critically short telomeres are more sensitive to 6-thio-dG and show a greater induction of telomere losses compared to cancer cells with long telomere reserves. Our studies reveal that telomere length and telomerase status determine 6-thio-dG sensitivity and uncover the molecular mechanism by which 6-thio-dG selectively inhibits telomerase synthesis of telomeric DNA.
Conflict of interest statement
DECLARATION OF INTERESTS The authors have no competing interests to declare.
Figures
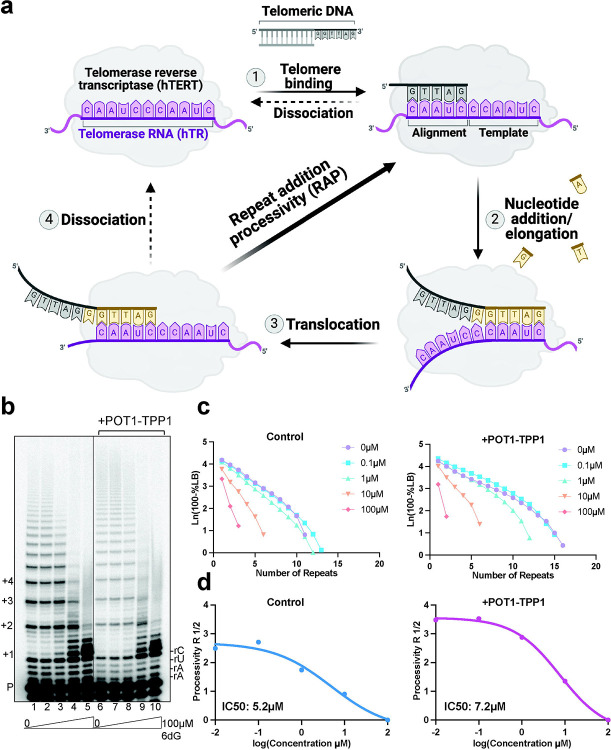
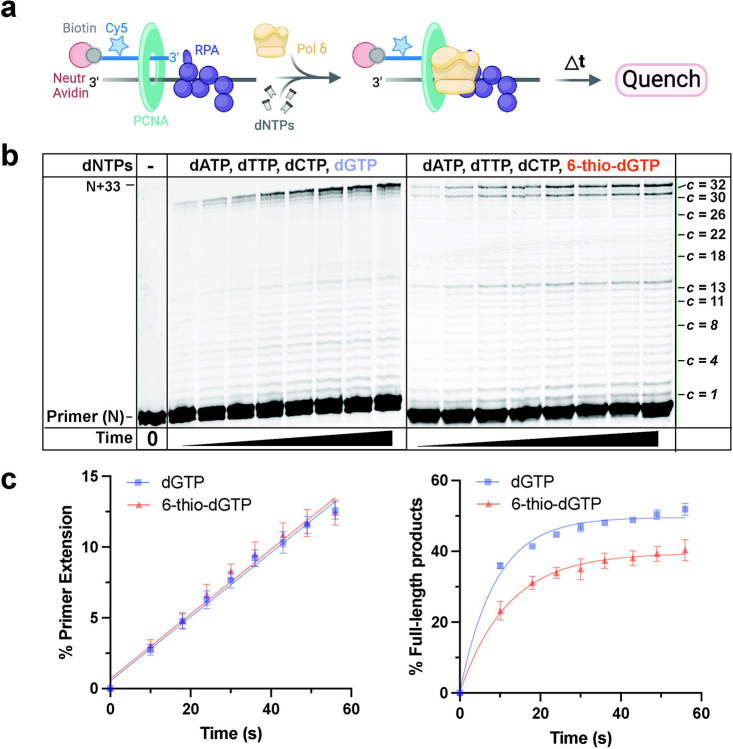
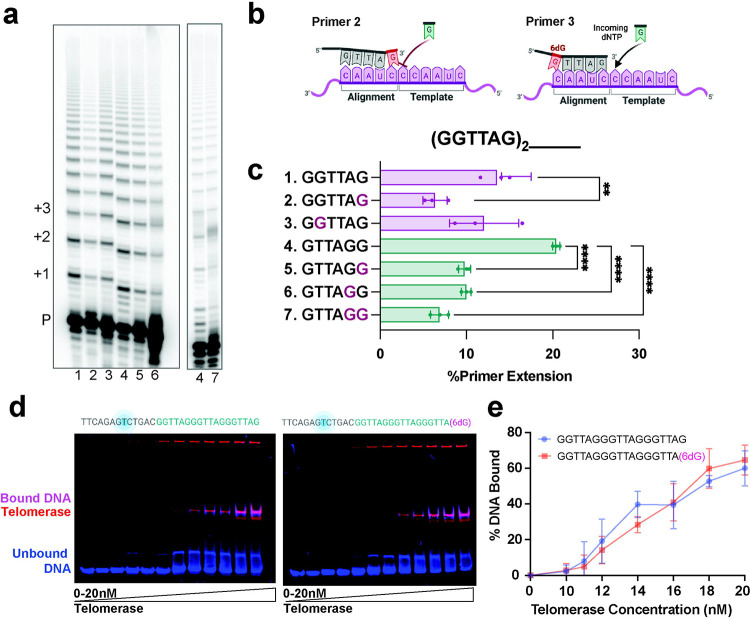
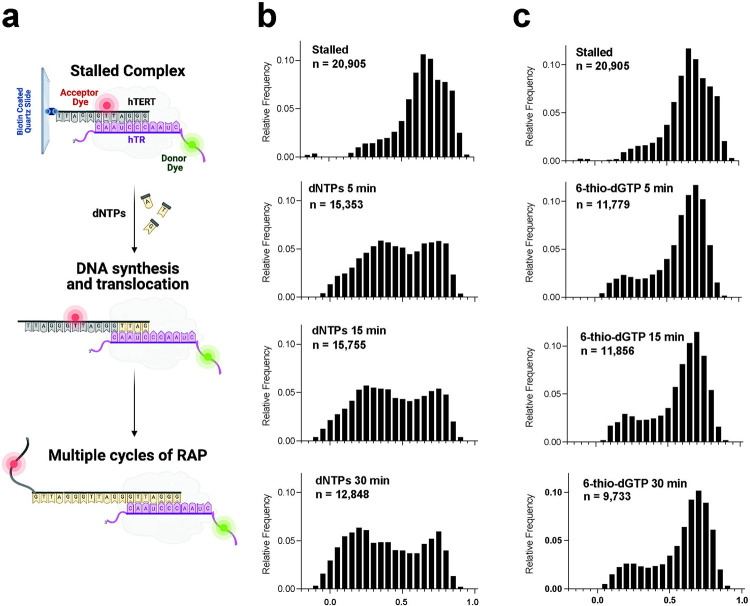
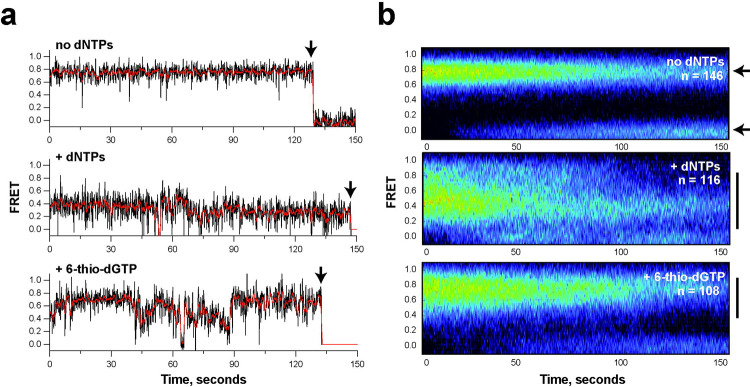
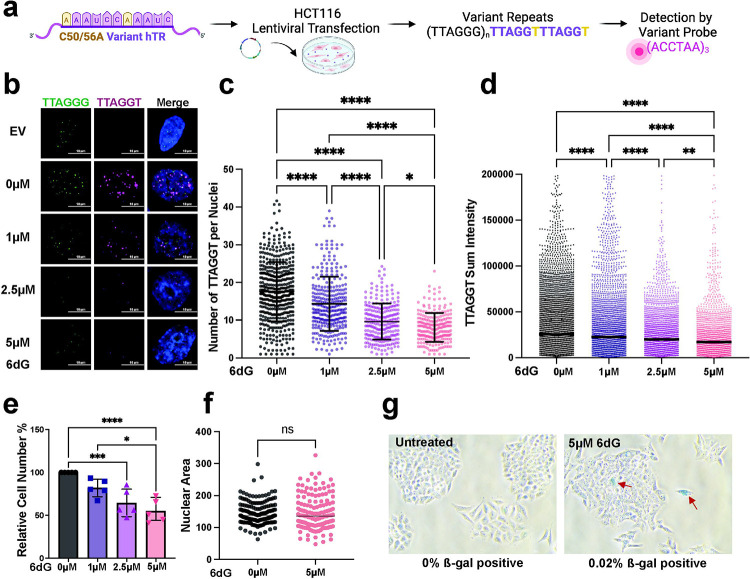
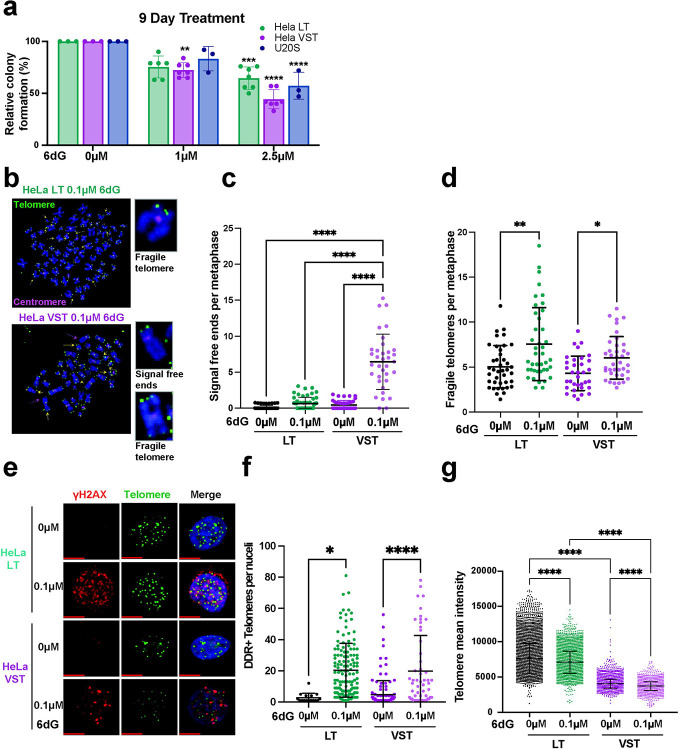
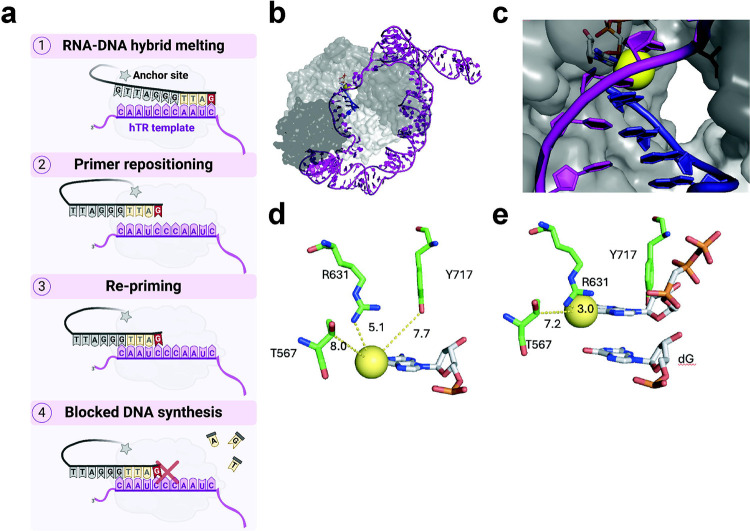
References
-
- de Lange T. Shelterin-Mediated Telomere Protection. Annu Rev Genet 52, 223–247 (2018). - PubMed
-
- d’Adda di Fagagna F. et al. A DNA damage checkpoint response in telomere-initiated senescence. Nature 426, 194–8 (2003). - PubMed
-
- Takai H., Smogorzewska A. & de Lange T. DNA damage foci at dysfunctional telomeres. Curr Biol 13, 1549–56 (2003). - PubMed
-
- Shay J.W. & Wright W.E. Senescence and immortalization: role of telomeres and telomerase. Carcinogenesis 26, 867–74 (2005). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources