

This is a preprint.
Structural basis for HIV-1 capsid adaption to rescue IP6-packaging deficiency
- PMID: 39975075
- PMCID: PMC11839029
- DOI: 10.1101/2025.02.09.637297
Structural basis for HIV-1 capsid adaption to rescue IP6-packaging deficiency
Update in
-
Structural basis for HIV-1 capsid adaption to a deficiency in IP6 packaging.Nat Commun. 2025 Sep 1;16(1):8152. doi: 10.1038/s41467-025-63363-9. Nat Commun. 2025. PMID: 40890096 Free PMC article.
Abstract
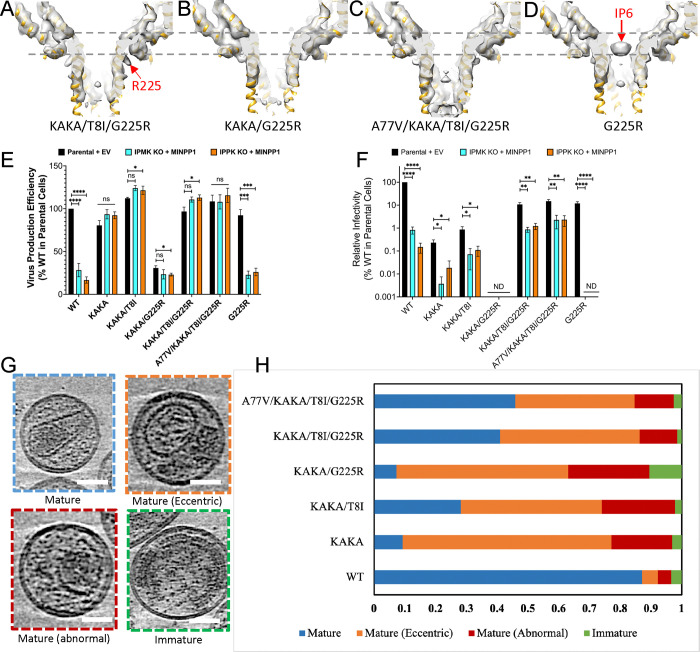
Inositol hexakisphosphate (IP6) promotes HIV-1 assembly via its interaction with the immature Gag lattice, effectively enriching IP6 within virions. During particle maturation, the HIV-1 protease cleaves the Gag polyproteins comprising the immature Gag lattice, releasing IP6 from its original binding site and liberating the capsid (CA) domain of Gag. IP6 then promotes the assembly of mature CA protein into the capsid shell of the viral core, which is required for infection of new target cells. Recently, we reported HIV-1 Gag mutants that assemble virions independently of IP6. However, these mutants are non-infectious and unable to assemble stable capsids. Here, we identified a mutation in the C-terminus of CA - G225R - that restores capsid formation and infectivity to these IP6-packaging-deficient mutants. Furthermore, we show that G225R facilitates the in vitro assembly of purified CA into capsid-like particles (CLPs) at IP6 concentrations well below those required for WT CLP assembly. Using single-particle cryoEM, we solved structures of CA hexamer and hexameric lattice of mature CLPs harbouring the G225R mutation assembled in low-IP6 conditions. The high-resolution (2.7 Å) cryoEM structure combined with molecular dynamics simulations of the G225R capsid revealed that the otherwise flexible and disordered C-terminus of CA becomes structured, extending to the pseudo two-fold hexamer-hexamer interface, thereby stabilizing the mature capsid. This work uncovers a structural mechanism by which HIV-1 adapts to a deficiency in IP6 packaging. Furthermore, the ability of G225R to promote mature capsid assembly in low-IP6 conditions provides a valuable tool for capsid-related studies and may indicate a heretofore unknown role for the unstructured C-terminus in HIV-1 capsid assembly.
Figures





References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous