

This is a preprint.
Ancient origin of an urban underground mosquito
- PMID: 39975080
- PMCID: PMC11838412
- DOI: 10.1101/2025.01.26.634793
Ancient origin of an urban underground mosquito
Update in
-
Ancient origin of an urban underground mosquito.Science. 2025 Oct 23;390(6771):eady4515. doi: 10.1126/science.ady4515. Epub 2025 Oct 23. Science. 2025. PMID: 41129649
Abstract
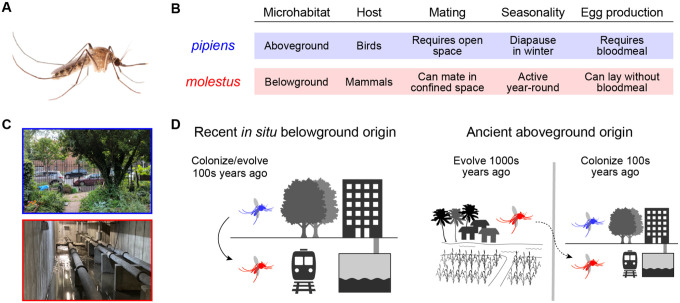
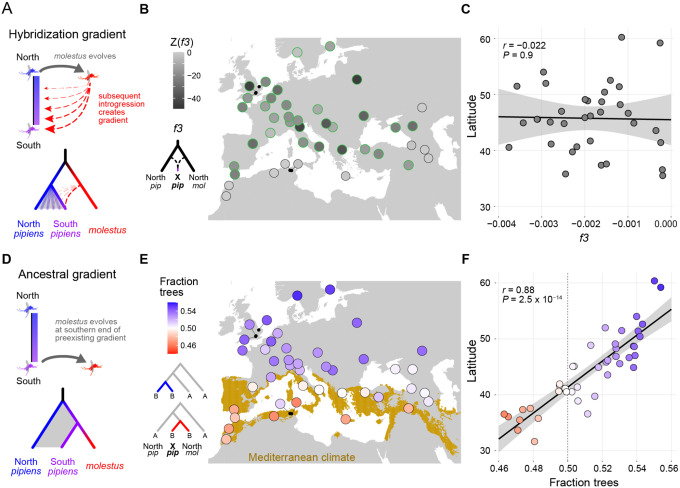
Understanding how life is adapting to urban environments represents an important challenge in evolutionary biology. Here we investigate a widely cited example of urban adaptation, Culex pipiens form molestus, also known as the London Underground Mosquito. Population genomic analysis of ~350 contemporary and historical samples counter the popular hypothesis that molestus originated belowground in London less than 200 years ago. Instead, we show that molestus first adapted to human environments aboveground in the Middle East over the course of >1000 years, likely in concert with the rise of agricultural civilizations. Our results highlight the role of early human society in priming taxa for contemporary urban evolution and have important implications for understanding arbovirus transmission.
Conflict of interest statement
Competing interests: None declared.
Figures




References
-
- Johnson M. T. J., Munshi-South J., Evolution of life in urban environments. Science 358, eaam8327 (2017). - PubMed
-
- Ritchie H., Samborska V., Roser M., Urbanization. Our world in data (2024).
-
- Vinogradova E. B., Culex Pipiens Pipiens Mosquitoes: Taxonomy, Distribution, Ecology, Physiology, Genetics, Applied Importance and Control (Pensoft, Sofia, Bulgaria, 2000)Pensoft series parasitologica.
-
- Harbach R., Culex pipiens: Species versus species complex – taxonomic history and perspective. J. Am. Mosq. Control Assoc. 28, 10–23 (2012). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources