

Multiple merger coalescent inference of effective population size
- PMID: 39976412
- PMCID: PMC11867189
- DOI: 10.1098/rstb.2023.0306
Multiple merger coalescent inference of effective population size
Abstract
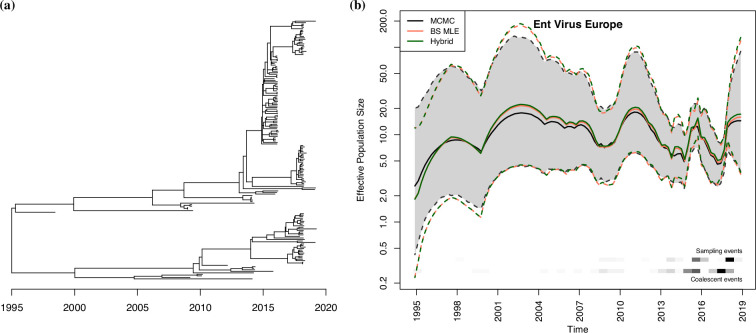
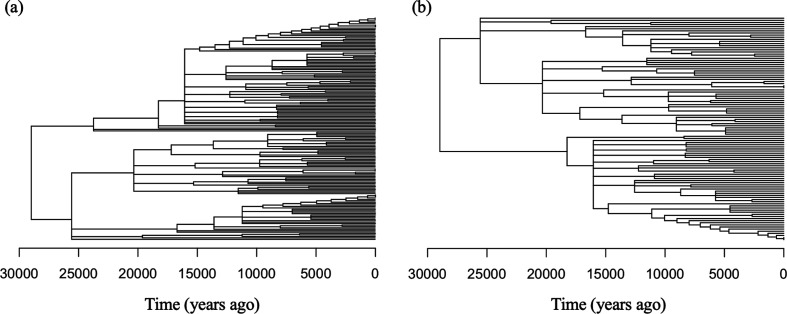
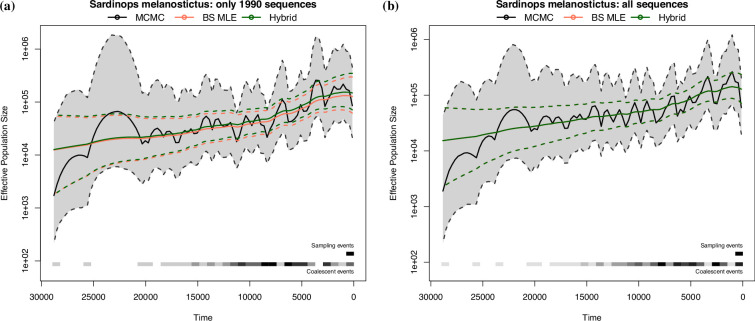
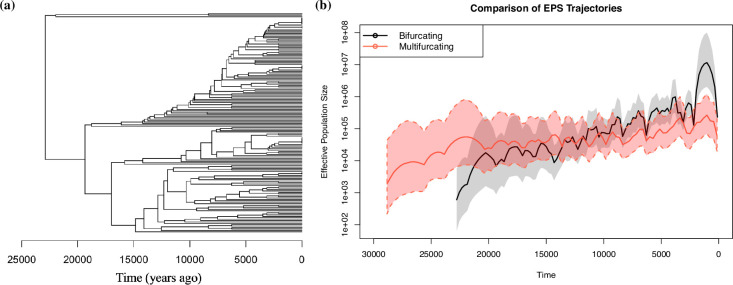
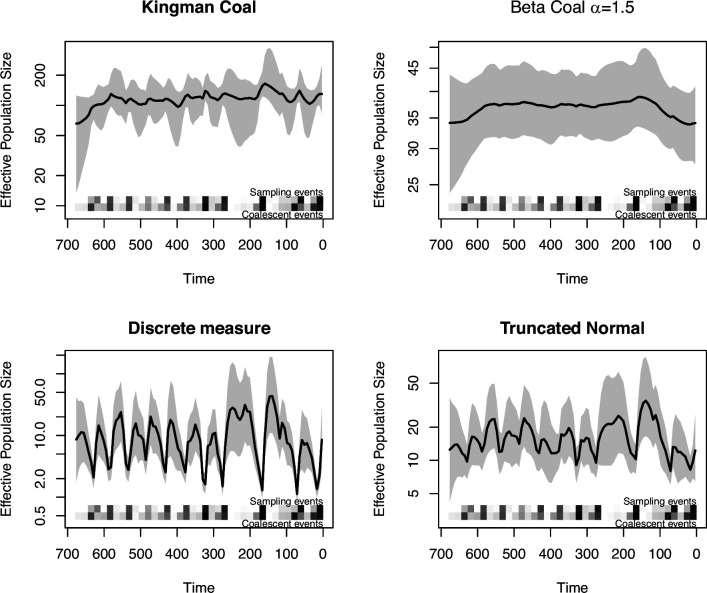
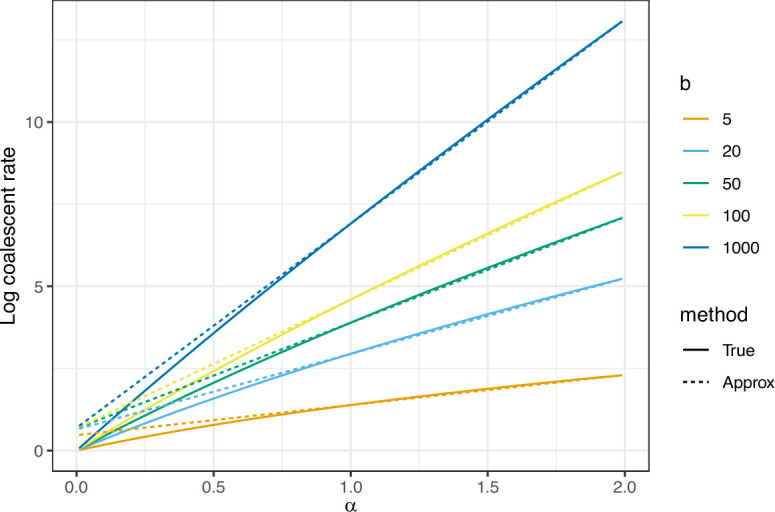
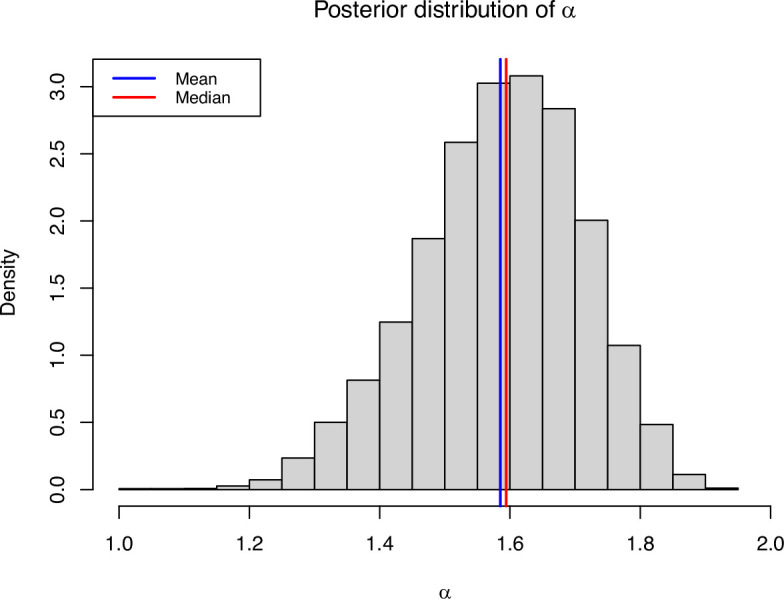
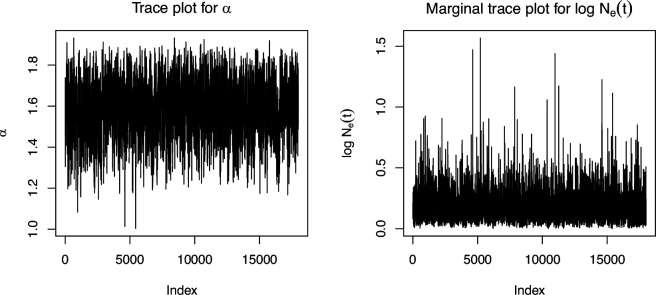
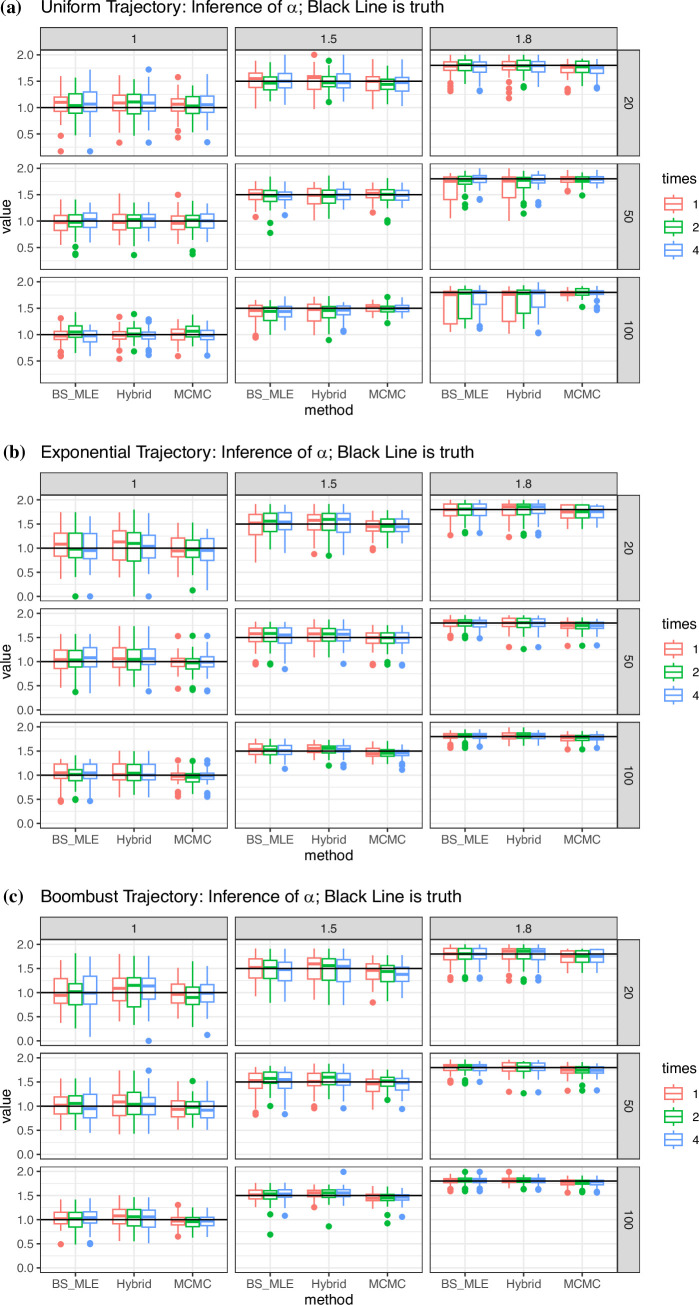
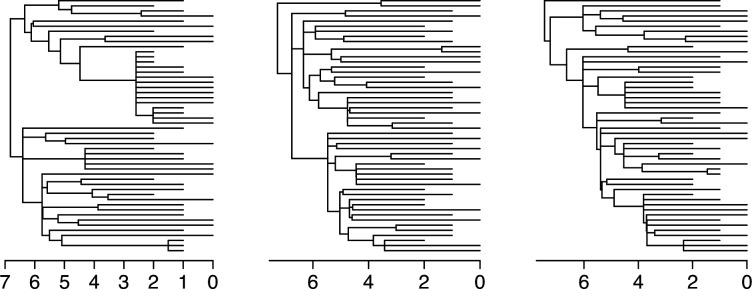
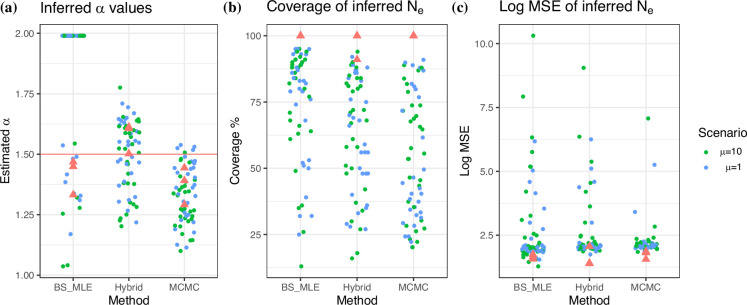
Variation in a sample of molecular sequence data informs about the past evolutionary history of the sample's population. Traditionally, Bayesian modelling coupled with the standard coalescent is used to infer the sample's bifurcating genealogy and demographic and evolutionary parameters such as effective population size and mutation rates. However, there are many situations where binary coalescent models do not accurately reflect the true underlying ancestral processes. Here, we propose a Bayesian non-parametric method for inferring effective population size trajectories from a multifurcating genealogy under the [Formula: see text]-coalescent. In particular, we jointly estimate the effective population size and the model parameter for the Beta-coalescent model, a special type of [Formula: see text]-coalescent. Finally, we test our methods on simulations and apply them to study various viral dynamics as well as Japanese sardine population size changes over time. The code and vignettes can be found in the phylodyn package.This article is part of the theme issue '"A mathematical theory of evolution": phylogenetic models dating back 100 years'.
Keywords: Beta-coalescent; Gaussian processes; Lambda-coalescent; Multiple mergers coalescent.
Conflict of interest statement
We declare we have no competing interests.
Figures
Similar articles
-
Inferring epidemiological dynamics with Bayesian coalescent inference: the merits of deterministic and stochastic models.Genetics. 2015 Feb;199(2):595-607. doi: 10.1534/genetics.114.172791. Epub 2014 Dec 19. Genetics. 2015. PMID: 25527289 Free PMC article.
-
Understanding Past Population Dynamics: Bayesian Coalescent-Based Modeling with Covariates.Syst Biol. 2016 Nov;65(6):1041-1056. doi: 10.1093/sysbio/syw050. Epub 2016 Jul 1. Syst Biol. 2016. PMID: 27368344 Free PMC article.
-
Bayesian Estimation of Population Size Changes by Sampling Tajima's Trees.Genetics. 2019 Nov;213(3):967-986. doi: 10.1534/genetics.119.302373. Epub 2019 Sep 11. Genetics. 2019. PMID: 31511299 Free PMC article.
-
Birth and death processes in phylogenetics and population genetics.Philos Trans R Soc Lond B Biol Sci. 2025 Feb 13;380(1919):20230300. doi: 10.1098/rstb.2023.0300. Epub 2025 Feb 20. Philos Trans R Soc Lond B Biol Sci. 2025. PMID: 39976404 Free PMC article. Review.
-
Coalescence 2.0: a multiple branching of recent theoretical developments and their applications.Mol Ecol. 2014 Jun;23(11):2637-52. doi: 10.1111/mec.12755. Epub 2014 May 20. Mol Ecol. 2014. PMID: 24750385 Review.
Cited by
-
"A mathematical theory of evolution": phylogenetic models dating back 100 years.Philos Trans R Soc Lond B Biol Sci. 2025 Feb 13;380(1919):20230297. doi: 10.1098/rstb.2023.0297. Epub 2025 Feb 20. Philos Trans R Soc Lond B Biol Sci. 2025. PMID: 39976405 Free PMC article. No abstract available.
References
-
- Kingman JFC. 1982. The coalescent. Stoch. Process. Their Appl. 13, 235–248. (10.1016/0304-4149(82)90011-4) - DOI
-
- Wakeley J. 2009. Coalescent theory: an introduction. Greenwood Village, CO: Roberts and Co.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
