

Increasingly efficient chromatin binding of cohesin and CTCF supports chromatin architecture formation during zebrafish embryogenesis
- PMID: 39979259
- PMCID: PMC11842872
- DOI: 10.1038/s41467-025-56889-5
Increasingly efficient chromatin binding of cohesin and CTCF supports chromatin architecture formation during zebrafish embryogenesis
Abstract
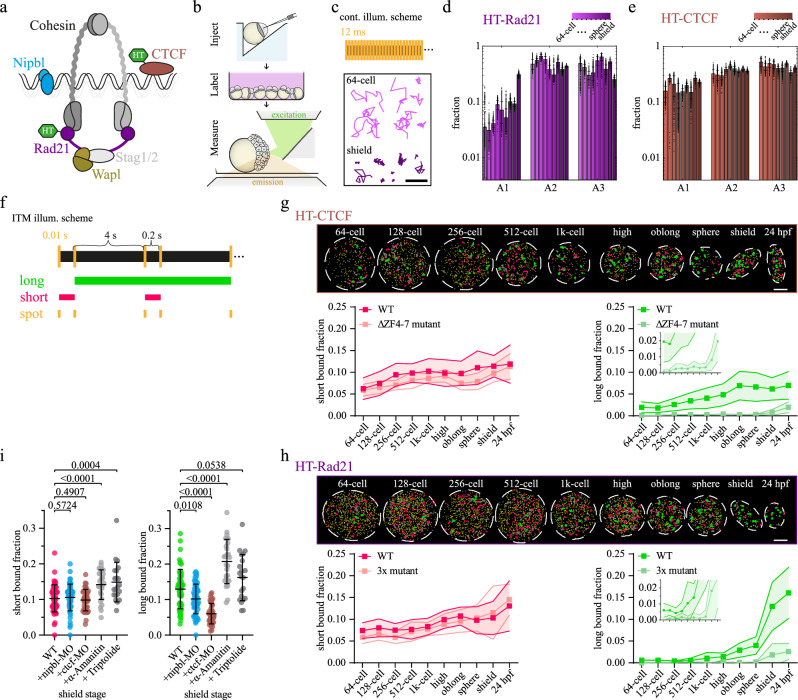
The three-dimensional folding of chromosomes is essential for nuclear functions such as DNA replication and gene regulation. The emergence of chromatin architecture is thus an important process during embryogenesis. To shed light on the molecular and kinetic underpinnings of chromatin architecture formation, we characterized biophysical properties of cohesin and CTCF binding to chromatin and their changes upon cofactor depletion using single-molecule imaging in live developing zebrafish embryos. We found that chromatin-bound fractions of both cohesin and CTCF increased significantly between the 1000-cell and shield stages, which we could explain through changes in both their association and dissociation rates. Moreover, increasing binding of cohesin restricted chromatin motion, potentially via loop extrusion, and showed distinct stage-dependent nuclear distribution. Polymer simulations with experimentally derived parameters recapitulated the experimentally observed gradual emergence of chromatin architecture. Our findings reveal molecular kinetics underlying chromatin architecture formation during zebrafish embryogenesis.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures



Similar articles
-
Cohesin facilitates zygotic genome activation in zebrafish.Development. 2018 Jan 3;145(1):dev156521. doi: 10.1242/dev.156521. Development. 2018. PMID: 29158440
-
CTCF and cohesin regulate chromatin loop stability with distinct dynamics.Elife. 2017 May 3;6:e25776. doi: 10.7554/eLife.25776. Elife. 2017. PMID: 28467304 Free PMC article.
-
Chromatin jets define the properties of cohesin-driven in vivo loop extrusion.Mol Cell. 2022 Oct 20;82(20):3769-3780.e5. doi: 10.1016/j.molcel.2022.09.003. Epub 2022 Sep 30. Mol Cell. 2022. PMID: 36182691
-
Beyond genomic weaving: molecular roles for CTCF outside cohesin loop extrusion.Curr Opin Genet Dev. 2025 Feb;90:102298. doi: 10.1016/j.gde.2024.102298. Epub 2024 Dec 21. Curr Opin Genet Dev. 2025. PMID: 39709822 Free PMC article. Review.
-
CTCF as a boundary factor for cohesin-mediated loop extrusion: evidence for a multi-step mechanism.Nucleus. 2020 Dec;11(1):132-148. doi: 10.1080/19491034.2020.1782024. Nucleus. 2020. PMID: 32631111 Free PMC article. Review.
Cited by
-
The physical chemistry of interphase loop extrusion.bioRxiv [Preprint]. 2024 Aug 24:2024.08.23.609419. doi: 10.1101/2024.08.23.609419. bioRxiv. 2024. PMID: 39229088 Free PMC article. Preprint.
-
Microscopy methods for the in vivo study of nanoscale nuclear organization.Biochem Soc Trans. 2025 Feb 3;53(1):BST20240629. doi: 10.1042/BST20240629. Biochem Soc Trans. 2025. PMID: 39898979 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
- 422780363/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 427512076/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 316249678/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 450627322/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 469281184/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 422389065/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 637987/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 European Research Council (H2020 Excellent Science - European Research Council)
- 759366/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 European Research Council (H2020 Excellent Science - European Research Council)
- 310030_192642/Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (Swiss National Science Foundation)
LinkOut - more resources
Full Text Sources
