

The polyamine transporter ATP13A3 mediates difluoromethylornithine-induced polyamine uptake in neuroblastoma
- PMID: 39981745
- PMCID: PMC11887671
- DOI: 10.1002/1878-0261.13789
The polyamine transporter ATP13A3 mediates difluoromethylornithine-induced polyamine uptake in neuroblastoma
Erratum in
-
Correction to "The polyamine transporter ATP13A3 mediates difluoromethylornithine-induced polyamine uptake in neuroblastoma".Mol Oncol. 2025 Aug;19(8):2457. doi: 10.1002/1878-0261.70088. Epub 2025 Jul 2. Mol Oncol. 2025. PMID: 40601788 Free PMC article. No abstract available.
Abstract
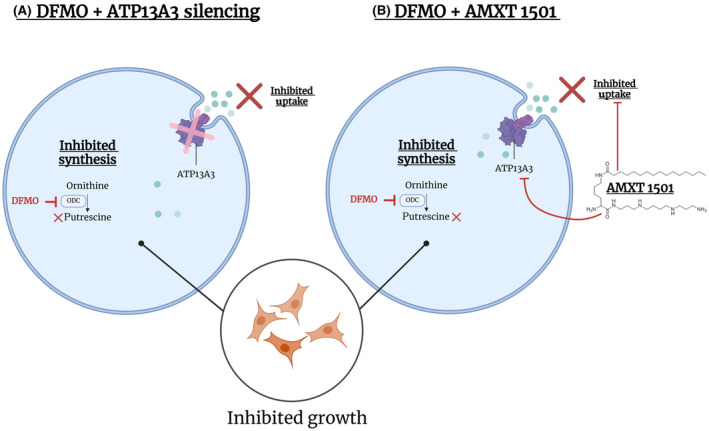
High-risk neuroblastomas, often associated with MYCN protooncogene amplification, are addicted to polyamines, small polycations vital for cellular functioning. We have previously shown that neuroblastoma cells increase polyamine uptake when exposed to the polyamine biosynthesis inhibitor difluoromethylornithine (DFMO), and this mechanism is thought to limit the efficacy of the drug in clinical trials. This finding resulted in the clinical development of polyamine transport inhibitors, including AMXT 1501, which is presently under clinical investigation in combination with DFMO. However, the mechanisms and transporters involved in DFMO-induced polyamine uptake are unknown. Here, we report that knockdown of ATPase 13A3 (ATP13A3), a member of the P5B-ATPase polyamine transporter family, limited basal and DFMO-induced polyamine uptake, attenuated MYCN-amplified and non-MYCN-amplified neuroblastoma cell growth, and potentiated the inhibitory effects of DFMO. Conversely, overexpression of ATP13A3 in neuroblastoma cells increased polyamine uptake, which was inhibited by AMXT 1501, highlighting ATP13A3 as a key target of the drug. An association between high ATP13A3 expression and poor survival in neuroblastoma further supports a role of this transporter in neuroblastoma progression. Thus, this study identified ATP13A3 as a critical regulator of basal and DFMO-induced polyamine uptake and a novel therapeutic target for neuroblastoma.
Keywords: AMXT 1501; ATP13A3; DFMO; neuroblastoma; polyamine depletion; polyamine transport inhibitor.
© 2025 Aminex Therapeutics, Inc. Children’s Cancer Institute and The Author(s). Molecular Oncology published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Conflict of interest statement
MRB is the Founder, President and CSO at Aminex Therapeutics where he is an employee and stock owner. PV is involved in polyamine transporter drug screening efforts for cancer and Parkinson's disease.
Figures









References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
