

Taming interleukin-12: Engineering of bispecific antibody-based IL-12 mimetics with biased agonism capacities
- PMID: 39981925
- PMCID: PMC11843475
- DOI: 10.1002/pro.70072
Taming interleukin-12: Engineering of bispecific antibody-based IL-12 mimetics with biased agonism capacities
Abstract
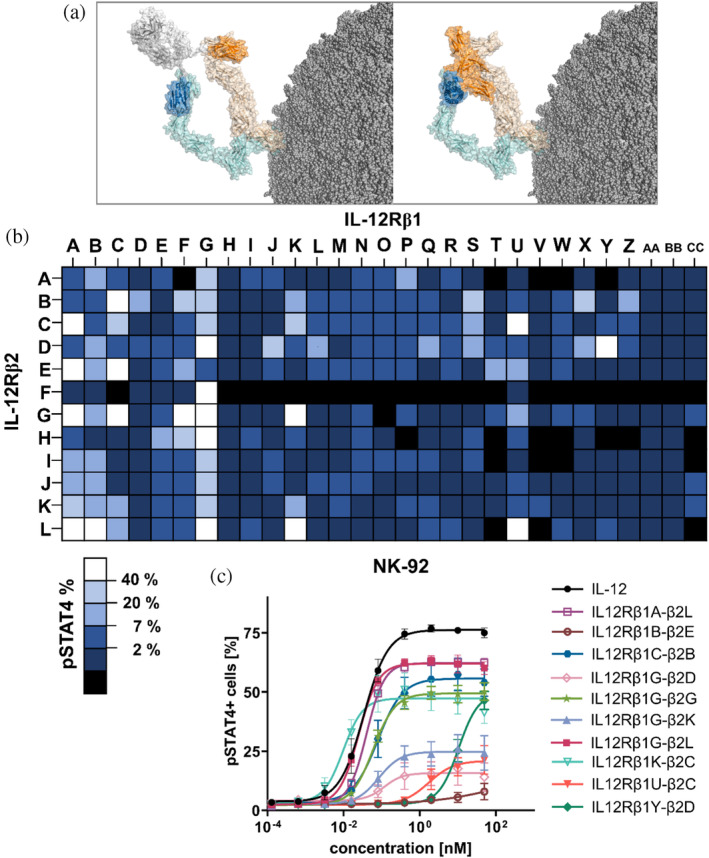
In this work, we have generated bispecific interleukin (IL)-12 surrogate agonists based on camelid-derived single-domain antibodies (sdAbs) targeting the IL-12 receptor (IL-12R) subunits IL-12Rβ1 and IL-12Rβ2. Following immunization and antibody display-based paratope isolation, respective sdAbs were combinatorially reformatted into a monovalent bispecific architecture by grafting resulting paratopes onto the hinge region of a heterodimeric Fc region. Functional characterization using NK-92 cells enabled the identification of multiple different sdAb-based bispecifics displaying divergent IL-12R agonism capacities as analyzed by STAT4 phosphorylation. Further investigations by harnessing peripheral blood mononuclear cells (PBMCs) from healthy donors revealed attenuated pSTAT4 activation compared to recombinant human (rh) wild-type IL-12 regarding both natural killer (NK)-cell and T-cell activation but robust IL-12R agonism on stimulated T cells. While several sdAb-based IL-12 mimetics were nearly inactive on NK cells as well as T cells obtained from PBMCs, they elicited significant STAT4 phosphorylation and interferon (IFN)-γ release on stimulated T cells as well as an IL-12-like transcriptional signature. Furthermore, we demonstrate that the activity of receptor agonism of generated bispecific IL-12 mimetics can also be biased towards stimulated T cells by changing the spatial orientation of the individual sdAbs within the molecular design architecture. Taken together, we present an alternative strategy to generate IL-12-like biologics with tailor-made characteristics.
Keywords: IL‐12; NK cell; T cell; VHH; antibody engineering; bispecific antibody; cytokine mimetic; single‐domain antibody; surrogate cytokine; yeast surface display.
© 2025 The Author(s). Protein Science published by Wiley Periodicals LLC on behalf of The Protein Society.
Conflict of interest statement
BL, LU, PA, SB, SK, JH, AD, LH, CX, AM, AE, DE, LP, and SZ are employees at either Merck Healthcare KGaA or EMD Serono. Besides, this work was conducted in the absence of any further commercial interest.
Figures



References
-
- Airoldi I, Gri G, Marshall JD, Corcione A, Facchetti P, Guglielmino R, et al. Expression and function of IL‐12 and IL‐18 receptors on human tonsillar B cells. J Immunol. 2000;165:6880–6888. - PubMed
-
- Bauer C, Ciesielski E, Pekar L, Krah S, Toleikis L, Zielonka S, et al. Facile one‐step generation of camelid VHH and avian scFv libraries for phage display by golden gate cloning. Methods Mol Biol. 2023;2681:47–60. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
