

Exploring the principles behind antibiotics with limited resistance
- PMID: 39984459
- PMCID: PMC11845477
- DOI: 10.1038/s41467-025-56934-3
Exploring the principles behind antibiotics with limited resistance
Abstract
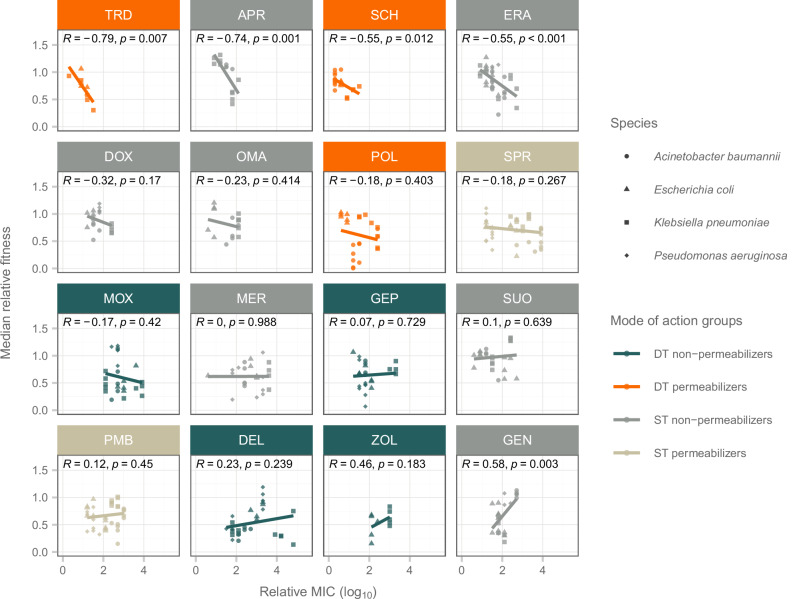
Antibiotics that target multiple cellular functions are anticipated to be less prone to bacterial resistance. Here we hypothesize that while dual targeting is crucial, it is not sufficient in preventing resistance. Only those antibiotics that simultaneously target membrane integrity and block another cellular pathway display reduced resistance development. To test the hypothesis, we focus on three antibiotic candidates, POL7306, Tridecaptin M152-P3 and SCH79797, all of which fulfill the above criteria. Here we show that resistance evolution against these antibiotics is limited in ESKAPE pathogens, including Escherichia coli, Klebsiella pneumoniae, Acinetobacter baumannii and Pseudomonas aeruginosa, while dual-target topoisomerase antibiotics are prone to resistance. We discover several mechanisms restricting resistance. First, de novo mutations result in only a limited elevation in resistance, including those affecting the molecular targets and efflux pumps. Second, resistance is inaccessible through gene amplification. Third, functional metagenomics reveal that mobile resistance genes are rare in human gut, soil and clinical microbiomes. Finally, we detect rapid eradication of bacterial populations upon toxic exposure to membrane targeting antibiotics. We conclude that resistance mechanisms commonly found in natural bacterial pathogens provide only limited protection to these antibiotics. Our work provides guidelines for the future development of antibiotics.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: Authors declare no competing interests.
Figures








References
-
- Lewis, K. The Science of Antibiotic Discovery. Cell181, 29–45 (2020). - PubMed
-
- Silver, L. L. Multi-targeting by monotherapeutic antibacterials. Nat. Rev. Drug Discov.6, 41–55 (2007). - PubMed
-
- Shukla, R. et al. An antibiotic from an uncultured bacterium binds to an immutable target. Cell186, 4059–4073.e27 (2023). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
