

Analysis of NS2-dependent effects on influenza PB1 segment extends replication requirements beyond the canonical promoter
- PMID: 39987189
- PMCID: PMC11846981
- DOI: 10.1038/s41467-025-57092-2
Analysis of NS2-dependent effects on influenza PB1 segment extends replication requirements beyond the canonical promoter
Abstract
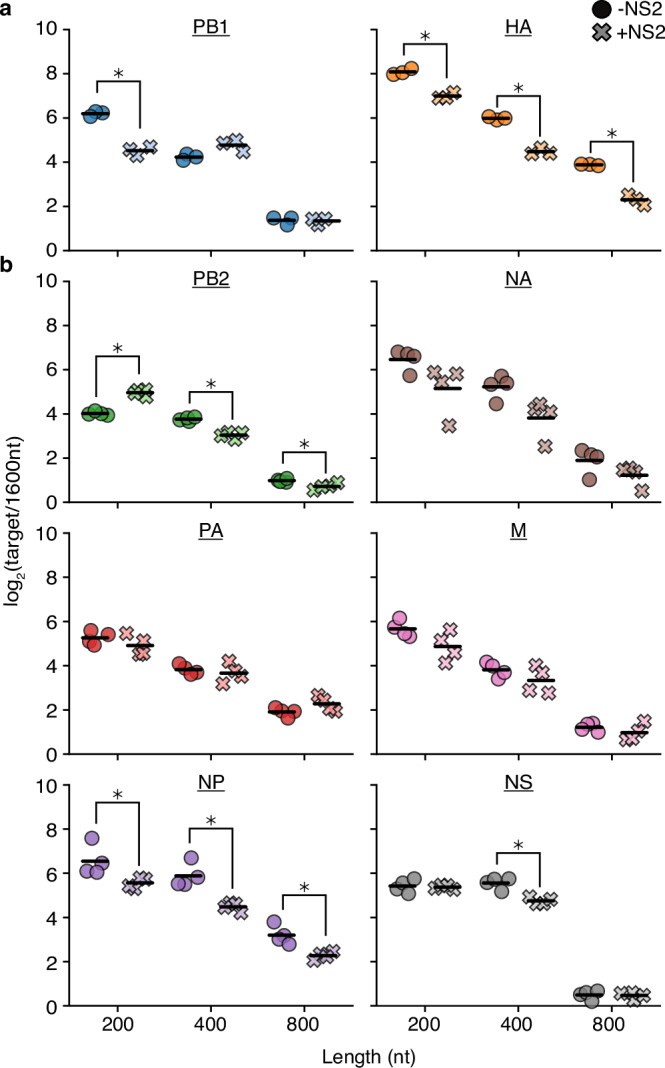
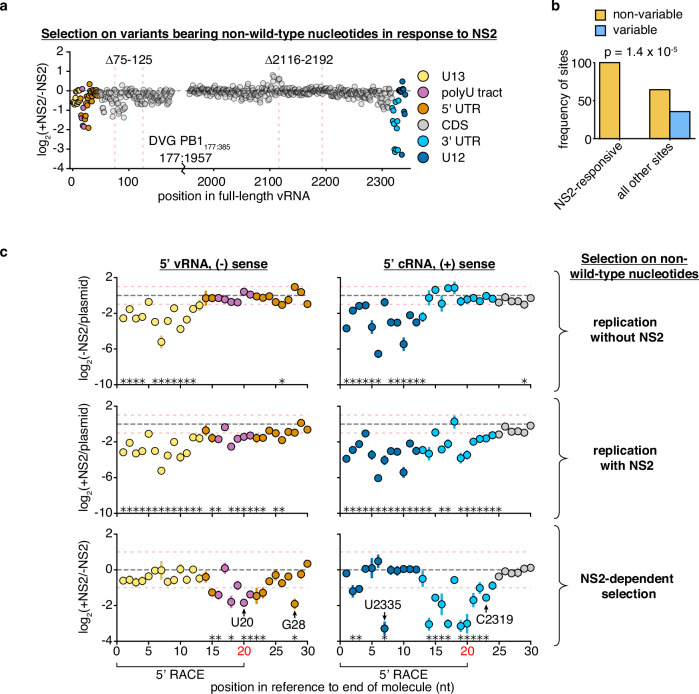
Influenza A virus encodes conserved promoter sequences. Using minimal replication assays-transfections with viral polymerase, nucleoprotein, and a genomic template-these sequences were identified as 13nt at the 5' end of the genomic RNA (U13) and 12nt at the 3' end (U12). Other than the fourth 3' nucleotide, the U12 and U13 sequences are identical between all eight RNA molecules of the segmented influenza A genome. However, individual segments can exhibit different dynamics during infection. Influenza NS2, which modulates transcription and replication differentially between genomic segments, may provide an explanation. Here, we assess how internal sequences of two genomic segments, HA and PB1, contribute to NS2-dependent replication and map such interactions down to individual nucleotides in PB1. We find that the expression of NS2 significantly alters sequence requirements for efficient replication beyond the identical U12 and U13 sequences, providing a potential mechanism for segment-specific replication dynamics across the influenza genome.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures




Update of
-
Efficient genome replication in influenza A virus requires NS2 and sequence beyond the canonical promoter.bioRxiv [Preprint]. 2024 Sep 10:2024.09.10.612348. doi: 10.1101/2024.09.10.612348. bioRxiv. 2024. Update in: Nat Commun. 2025 Feb 22;16(1):1875. doi: 10.1038/s41467-025-57092-2. PMID: 39314307 Free PMC article. Updated. Preprint.
References
-
- Robb, N. C., Smith, M., Vreede, F. T. & Fodor, E. NS2/NEP protein regulates transcription and replication of the influenza virus RNA genome. J. Gen. Virol.90, 1398–1407 (2009). - PubMed
MeSH terms
Substances
Grants and funding
- 1T32GM148739-01A1/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- DGE-2039656/National Science Foundation (NSF)
- T32 GM148739/GM/NIGMS NIH HHS/United States
- R35GM147031/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- DP2 AI175474/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Miscellaneous
