

Structural basis of endo-siRNA processing by Drosophila Dicer-2 and Loqs-PD
- PMID: 39988314
- PMCID: PMC11840564
- DOI: 10.1093/nar/gkaf102
Structural basis of endo-siRNA processing by Drosophila Dicer-2 and Loqs-PD
Abstract
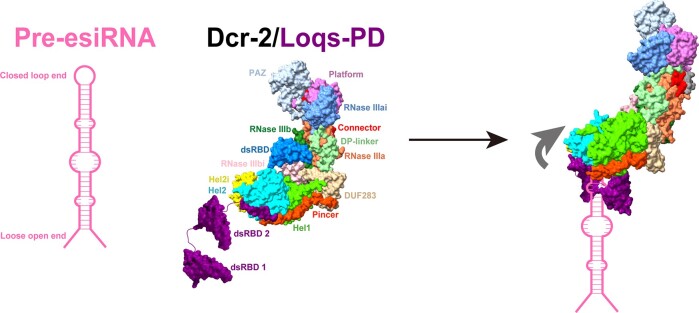
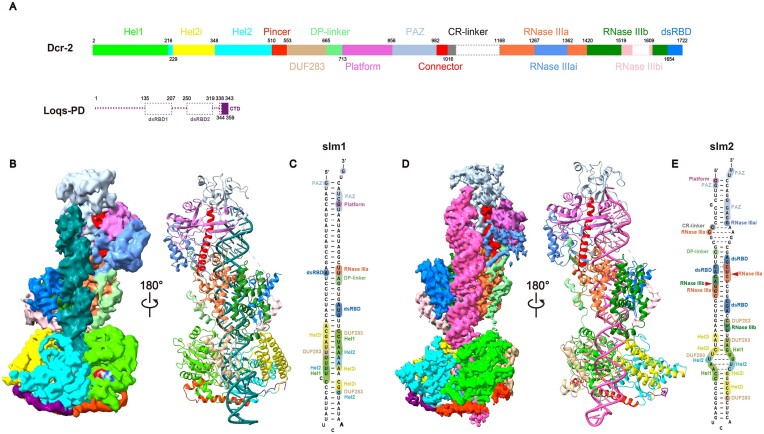
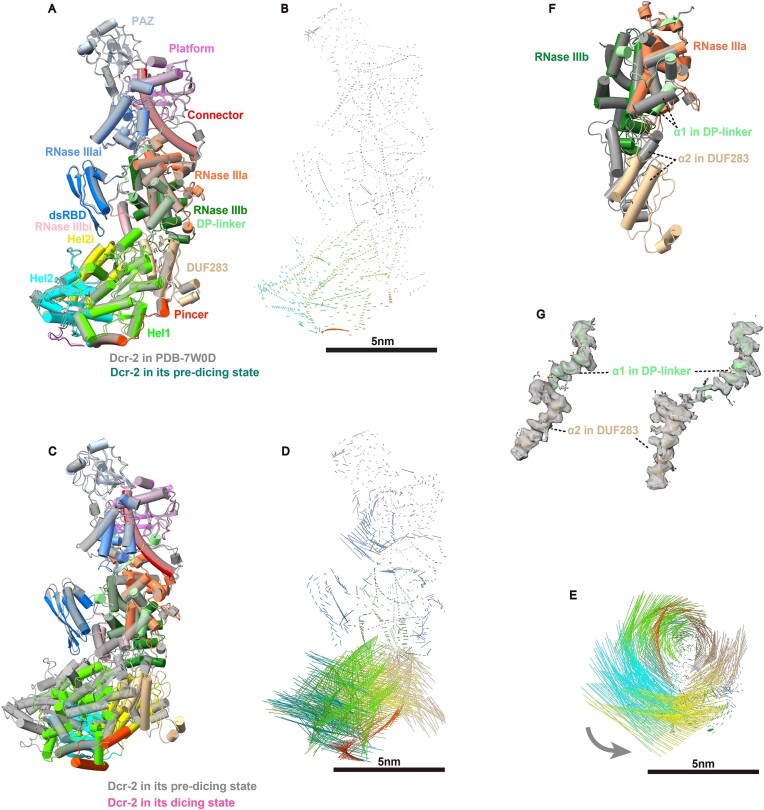
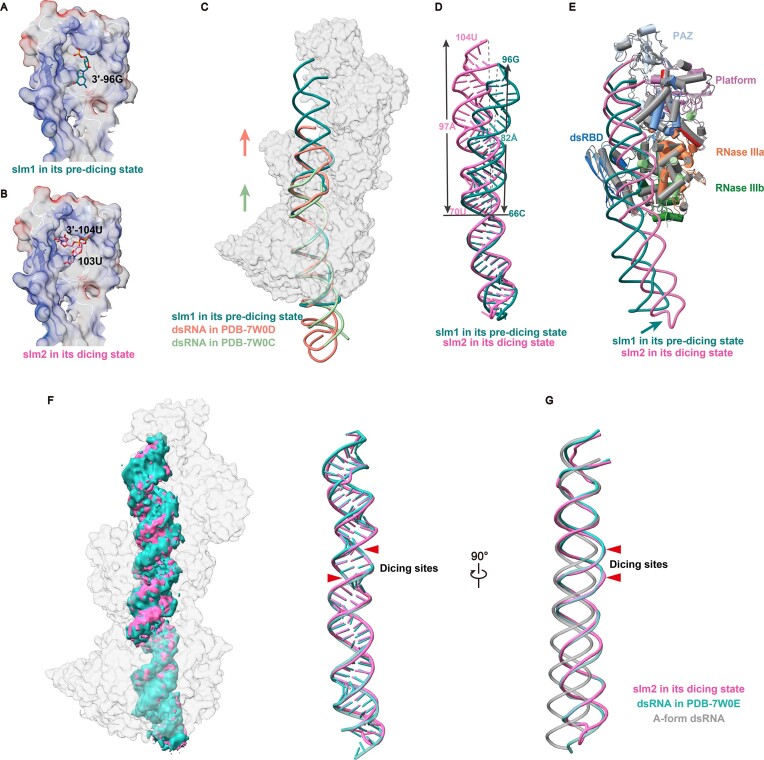
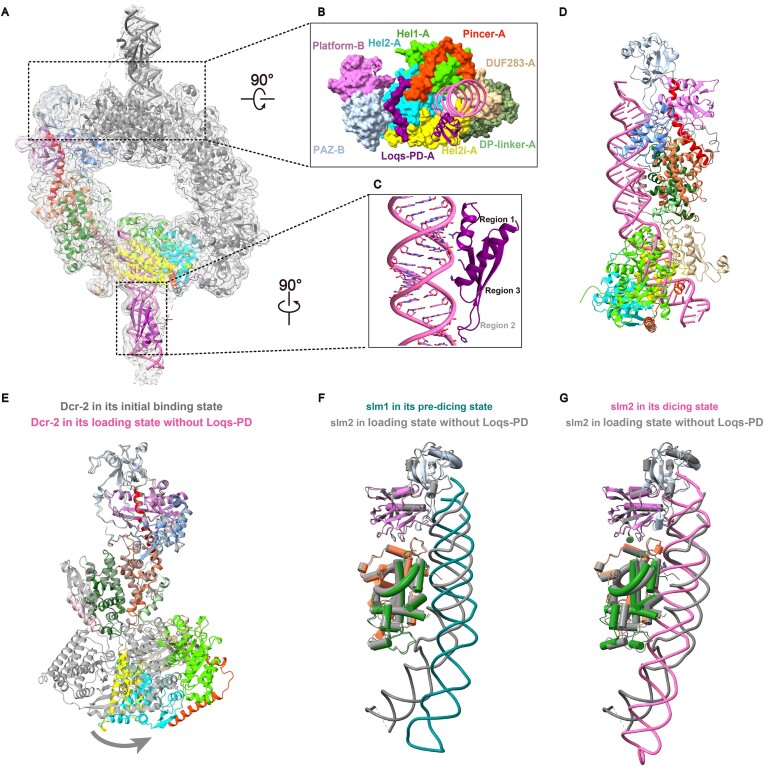
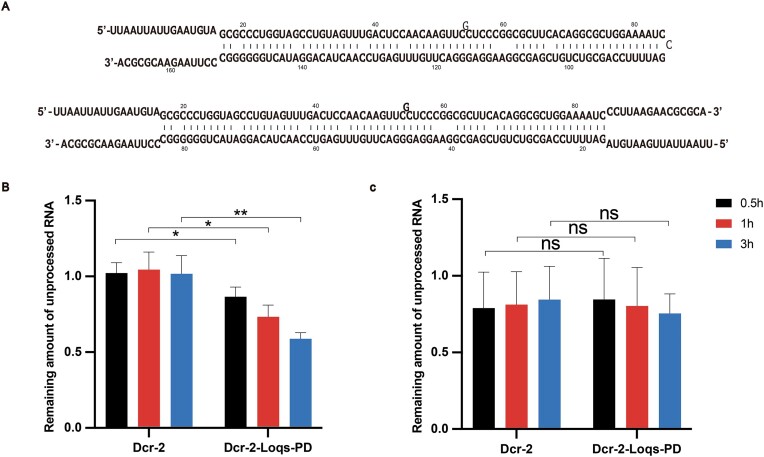
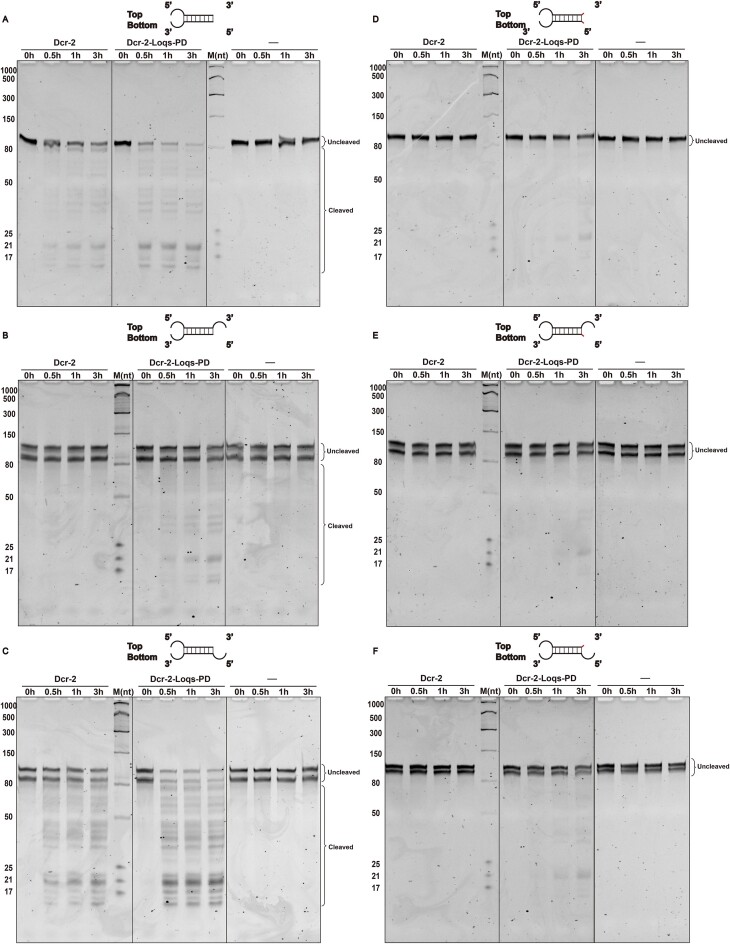
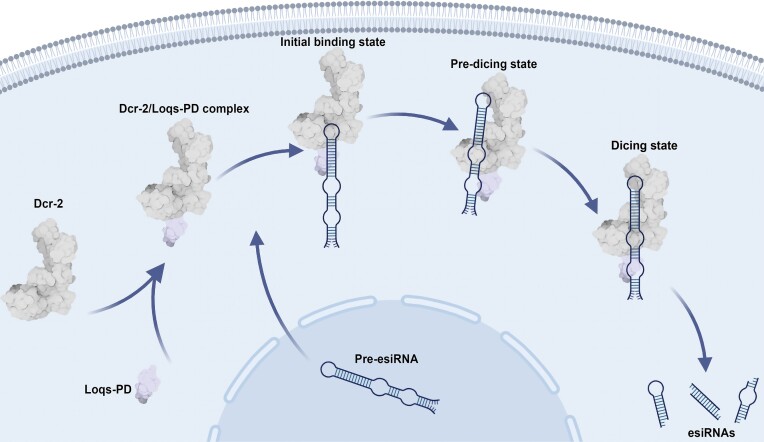
Endogenous small interfering RNAs (endo-siRNAs or esiRNAs) originate from either elongated endogenous transcripts capable of forming complex fold-back structures or from double-stranded regions generated through intermolecular base pairing of convergently transcribed mRNAs. The mechanism of maturation and functionality of esiRNAs exhibit significant variation across diverse species. In Drosophila melanogaster, esiRNAs reside in both somatic and germline cells, where they serve as post-transcriptional modulators for specific target RNAs. Their maturation process critically relies on Dicer-2 (Dcr-2), with the assistance of its cofactor Loqs-PD. In this study, we have successfully elucidated the cryo-EM structures of Dcr-2/Loqs-PD complex bound to esiRNA precursors (pre-esiRNAs) in various states. Our structural and biochemical results reveal that ATP is essential for the cleavage of esiRNAs by the Dcr-2/Loqs-PD complex, a process analogous to the cleavage of double-stranded RNA (dsRNA). When Loqs-PD is present, pre-esiRNAs are preferentially loaded onto the Helicase domain of Dcr-2. Moreover, as the Helicase domain exhibits a preference for binding to the rigid end of double-stranded RNA, Dcr-2 tends to cleave pre-esiRNA from the small closed loop end, rather than the loose and flexible open end.
© The Author(s) 2025. Published by Oxford University Press on behalf of Nucleic Acids Research.
Conflict of interest statement
None declared.
Figures
Similar articles
-
Structural insights into dsRNA processing by Drosophila Dicer-2-Loqs-PD.Nature. 2022 Jul;607(7918):399-406. doi: 10.1038/s41586-022-04911-x. Epub 2022 Jun 29. Nature. 2022. PMID: 35768513 Free PMC article.
-
Loquacious-PD facilitates Drosophila Dicer-2 cleavage through interactions with the helicase domain and dsRNA.Proc Natl Acad Sci U S A. 2017 Sep 19;114(38):E7939-E7948. doi: 10.1073/pnas.1707063114. Epub 2017 Sep 5. Proc Natl Acad Sci U S A. 2017. PMID: 28874570 Free PMC article.
-
Molecular mechanisms that funnel RNA precursors into endogenous small-interfering RNA and microRNA biogenesis pathways in Drosophila.RNA. 2010 Mar;16(3):506-15. doi: 10.1261/rna.1952110. Epub 2010 Jan 19. RNA. 2010. PMID: 20086050 Free PMC article.
-
Cryo-EM structures of human DICER dicing a pre-miRNA substrate.FEBS J. 2024 Jul;291(14):3072-3079. doi: 10.1111/febs.17048. Epub 2024 Jan 10. FEBS J. 2024. PMID: 38151772 Review.
-
The RNAi pathway initiated by Dicer-2 in Drosophila.Cold Spring Harb Symp Quant Biol. 2006;71:39-44. doi: 10.1101/sqb.2006.71.008. Cold Spring Harb Symp Quant Biol. 2006. PMID: 17381278 Review.
Cited by
-
Biochemical and structural basis of Dicer helicase function unveiled by resurrecting ancient proteins.Proc Natl Acad Sci U S A. 2025 Jun 3;122(22):e2500825122. doi: 10.1073/pnas.2500825122. Epub 2025 May 28. Proc Natl Acad Sci U S A. 2025. PMID: 40434637
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
