

This is a preprint.
A connection between Vibrio cholerae motility and inter-animal transmission
- PMID: 39990368
- PMCID: PMC11844489
- DOI: 10.1101/2025.02.12.637895
A connection between Vibrio cholerae motility and inter-animal transmission
Update in
-
Vibrio cholerae motility is associated with inter-animal transmission.Nat Commun. 2025 Aug 27;16(1):7989. doi: 10.1038/s41467-025-62984-4. Nat Commun. 2025. PMID: 40866349 Free PMC article.
Abstract
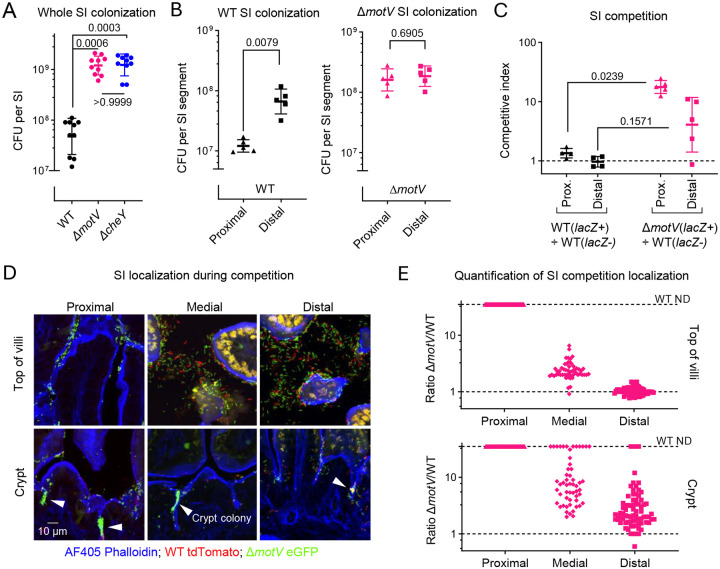
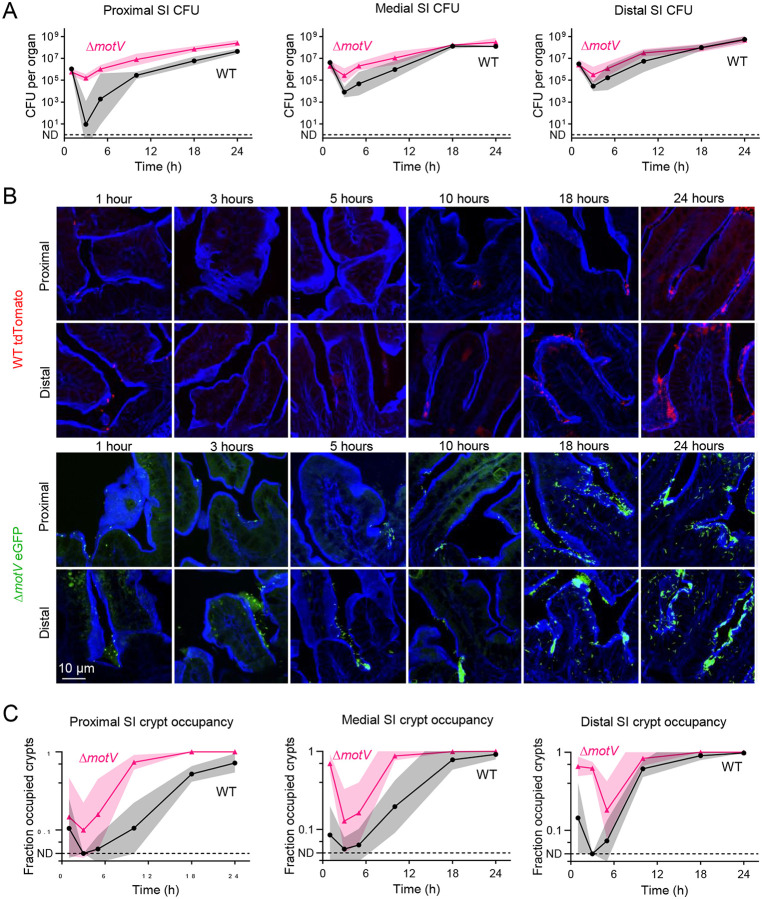
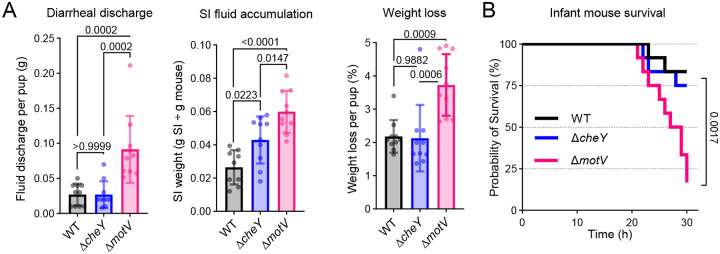
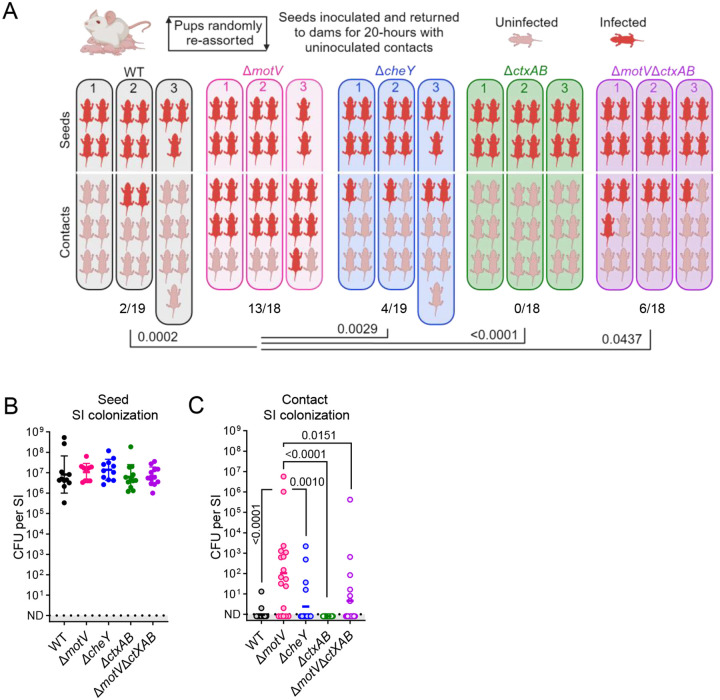
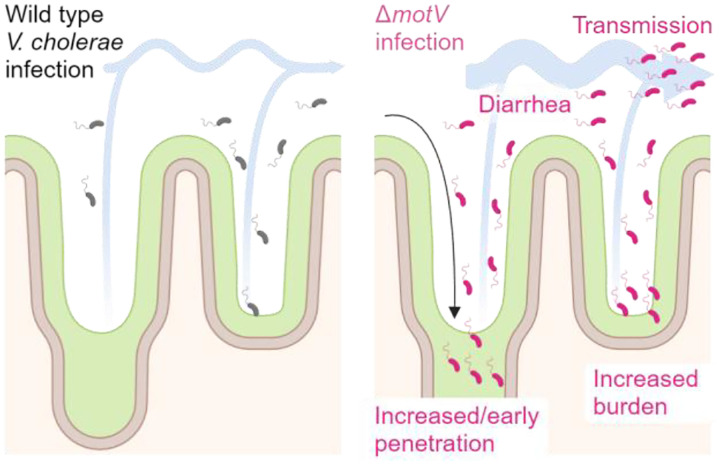
Outbreaks of cholera are caused by the highly transmissive pathogen Vibrio cholerae. Here, a transposon screen revealed that inactivation of the V. cholerae motility-linked gene motV increases infant mouse intestinal colonization. Compared to wild-type V. cholerae, a ΔmotV mutant, which exhibits heightened motility in the form of constitutive straight swimming, localizes to the crypts earlier in infection and over a larger area of the small intestine. Aberrant localization of the mutant was associated with an increased number of V. cholerae initiating infection, and elevated pathogen burden, diarrhea, and lethality. Moreover, the deletion of motV causes V. cholerae to transmit from infected suckling mice to naïve littermates more efficiently. Even in the absence of cholera toxin, the ΔmotV mutant continues to transmit between animals, although less than in the presence of toxin, indicating that phenotypes other than cholera toxin-driven diarrhea contribute to transmission. Collectively, this work provides experimental evidence linking intra-animal bottlenecks, colonization, and disease to inter-animal transmission.
Conflict of interest statement
Competing Interests The authors declare no competing interests.
Figures


References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources