

The structure of basal body inner junctions from Tetrahymena revealed by electron cryo-tomography
- PMID: 39994484
- PMCID: PMC11961760
- DOI: 10.1038/s44318-025-00392-6
The structure of basal body inner junctions from Tetrahymena revealed by electron cryo-tomography
Abstract
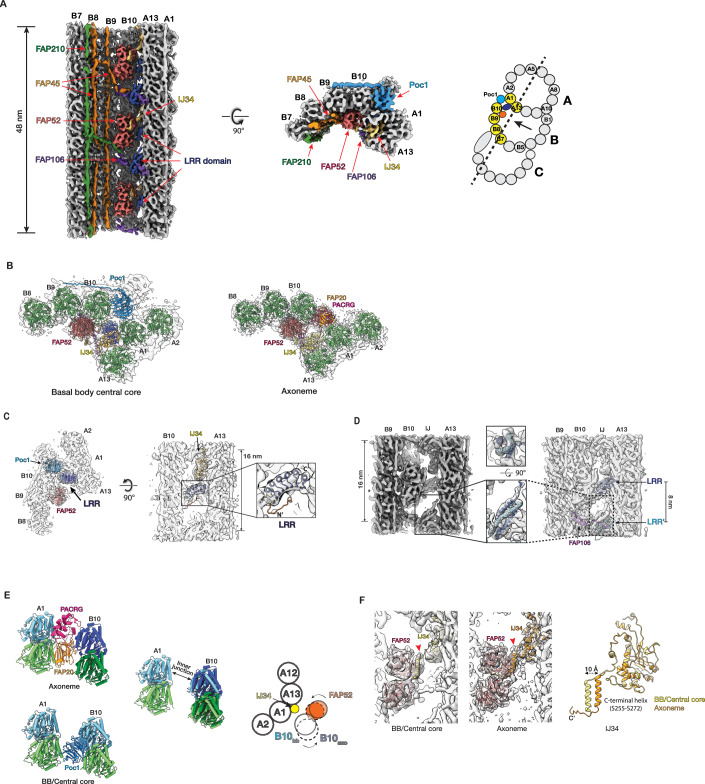
The cilium is a microtubule-based eukaryotic organelle critical for many cellular functions. Its assembly initiates at a basal body and continues as an axoneme that projects out of the cell to form a functional cilium. This assembly process is tightly regulated. However, our knowledge of the molecular architecture and the mechanism of assembly is limited. By applying cryo-electron tomography, we obtained structures of the inner junction in three regions of the cilium from Tetrahymena: the proximal, the central core of the basal body, and the axoneme. We identified several protein components in the basal body. While a few proteins are distributed throughout the entire length of the organelle, many are restricted to specific regions, forming intricate local interaction networks in the inner junction and bolstering local structural stability. By examining the inner junction in a POC1 knockout mutant, we found the triplet microtubule was destabilized, resulting in a defective structure. Surprisingly, several axoneme-specific components were found to "infiltrate" into the mutant basal body. Our findings provide molecular insight into cilium assembly at the inner junctions, underscoring its precise spatial regulation.
Keywords: Assembly; Basal Body; Centriole; Cilium; Electron Cryo-tomography.
© 2025. The Author(s).
Conflict of interest statement
Disclosure and competing interests statement. The authors declare no competing interests.
Figures










Update of
-
The Structure of Cilium Inner Junctions Revealed by Electron Cryo-tomography.bioRxiv [Preprint]. 2024 Sep 9:2024.09.09.612100. doi: 10.1101/2024.09.09.612100. bioRxiv. 2024. Update in: EMBO J. 2025 Apr;44(7):1975-2001. doi: 10.1038/s44318-025-00392-6. PMID: 39314311 Free PMC article. Updated. Preprint.
References
-
- Breugel M, van, Hirono M, Andreeva A, Yanagisawa H, Yamaguchi S, Nakazawa Y, Morgner N, Petrovich M, Ebong I-O, Robinson CV et al (2011) Structures of SAS-6 Suggest Its Organization in Centrioles. Science 331:1196–1199 - PubMed
