

Echinocandin Adaptation in Candida albicans Is Accompanied by Altered Chromatin Accessibility at Gene Promoters and by Cell Wall Remodeling
- PMID: 39997404
- PMCID: PMC11856910
- DOI: 10.3390/jof11020110
Echinocandin Adaptation in Candida albicans Is Accompanied by Altered Chromatin Accessibility at Gene Promoters and by Cell Wall Remodeling
Abstract
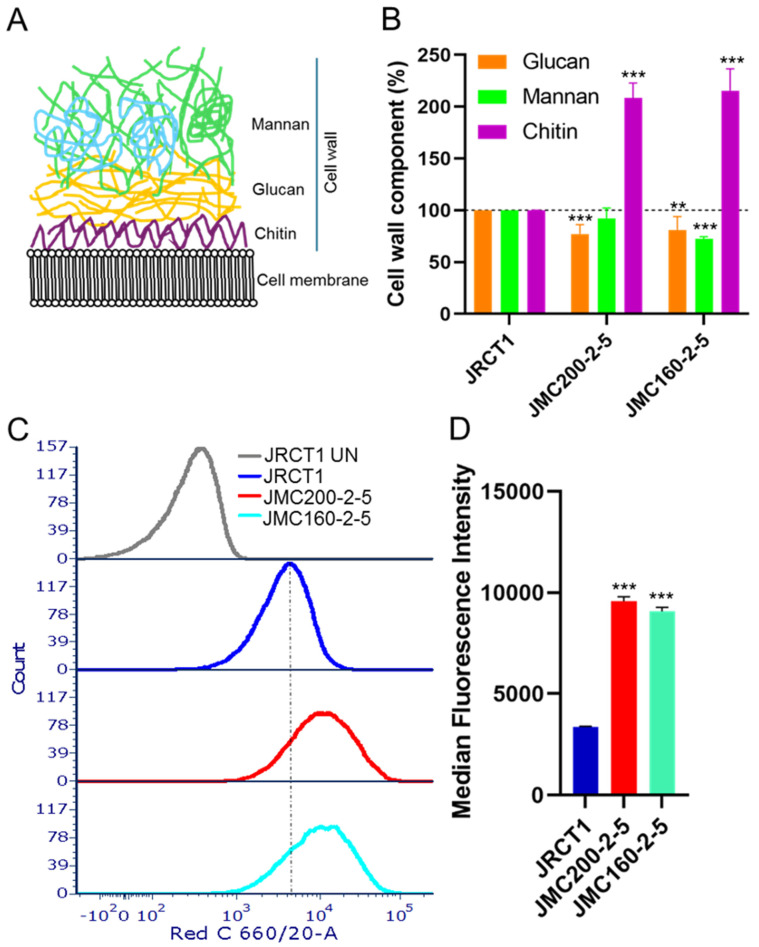
Infections by the major opportunistic pathogen of human Candida albicans are commonly treated with echinocandin (ECN) drugs. However, C. albicans can adapt to grow in the presence of certain amounts of ECNs. Prior studies by several laboratories have defined multiple genes, as well as mechanisms involving induced aneuploidy, that can govern this. Still, the mechanisms of ECN adaptation are not fully understood. Here, we use genome-wide profiling of chromatin accessibility by ATAC-seq to determine if ECN adaptation is reflected in changes in the chromatin landscape in the absence of aneuploidy. We find that drug adaptation is coupled with multiple changes in chromatin accessibility genome-wide, which occur predominantly in gene promoter regions. Areas of increased accessibilities in promoters are enriched with the binding motifs for at least two types of transcription factors: zinc finger and basic leucine zipper. We also find that chromatin changes are often associated with differentially expressed genes including genes with functions relevant to the ECN-adapted phenotype, such as cell wall biosynthesis. Consistent with this, we find that the cell wall is remodeled in ECN-adapted mutants, with chitin up and glucan down and increased cell surface exposure. A full understanding of ECN adaptation processes is of critical importance for the prevention of clinical resistance.
Keywords: ATAC-seq; Candida albicans; echinocandin adaptation.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures







Similar articles
-
At least 10 genes on chromosome 5 of Candida albicans are downregulated in concert to control cell wall and to confer adaptation to caspofungin.bioRxiv [Preprint]. 2023 Feb 6:2023.02.06.527048. doi: 10.1101/2023.02.06.527048. bioRxiv. 2023. PMID: 36798197 Free PMC article. Preprint.
-
Genome-Wide DNA Changes Acquired by Candida albicans Caspofungin-Adapted Mutants.Microorganisms. 2023 Jul 25;11(8):1870. doi: 10.3390/microorganisms11081870. Microorganisms. 2023. PMID: 37630430 Free PMC article.
-
Candida albicans Genes Modulating Echinocandin Susceptibility of Caspofungin-Adapted Mutants Are Constitutively Expressed in Clinical Isolates with Intermediate or Full Resistance to Echinocandins.J Fungi (Basel). 2024 Mar 19;10(3):224. doi: 10.3390/jof10030224. J Fungi (Basel). 2024. PMID: 38535232 Free PMC article.
-
Candida and candidaemia. Susceptibility and epidemiology.Dan Med J. 2013 Nov;60(11):B4698. Dan Med J. 2013. PMID: 24192246 Review.
-
Echinocandin resistance, susceptibility testing and prophylaxis: implications for patient management.Drugs. 2014 Sep;74(14):1573-85. doi: 10.1007/s40265-014-0286-5. Drugs. 2014. PMID: 25255923 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
