

Atypical memory B cells acquire Breg phenotypes in hepatocellular carcinoma
- PMID: 39998891
- PMCID: PMC11981623
- DOI: 10.1172/jci.insight.187025
Atypical memory B cells acquire Breg phenotypes in hepatocellular carcinoma
Abstract
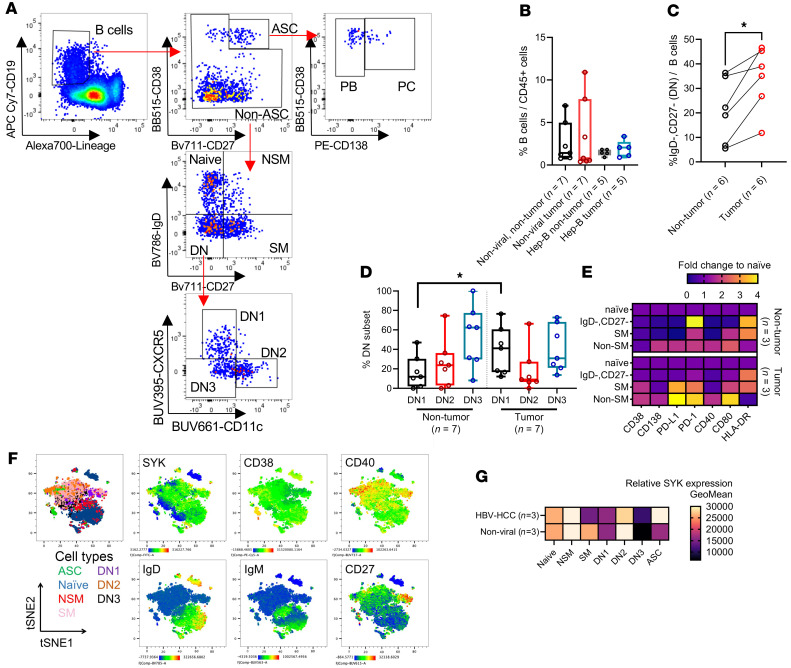
The functional plasticity of tumor-infiltrating lymphocyte B-cells (TIL-B) spans from antitumor responses to noncanonical immune suppression. Yet, how the tumor microenvironment (TME) influences TIL-B development is still underappreciated. Our current study integrated single-cell transcriptomics and B cell receptor (BCR) sequencing to profile TIL-B phenotypes and clonalities in hepatocellular carcinoma (HCC). Using trajectory and gene regulatory network analysis, we were able to characterize plasma cells and memory and naive B cells within the HCC TME and further revealed a downregulation of BCR signaling genes in plasma cells and a subset of inflammatory TNF+ memory B cells. Within the TME, a nonswitched memory B cell subset acquired an age-associated B cell phenotype (TBET+CD11c+) and expressed higher levels of PD-L1, CD25, and granzyme B. We further demonstrated that the presence of HCC tumor cells could confer suppressive functions on peripheral blood B cells that in turn, dampen T cell costimulation. To the best of our knowledge, these findings represent novel mechanisms of noncanonical immune suppression in HCC. While previous studies identified atypical memory B cells in chronic hepatitis and across several solid cancer types, we further highlighted their potential role as regulatory B cells (Bregs) within both the TME and peripheral blood of HCC patients.
Keywords: Adaptive immunity; Hepatology; Immunology.
Conflict of interest statement
Figures





References
-
- Xiao X, et al. PD-1hi identifies a novel regulatory B-cell population in human hepatoma that promotes disease progression. Cancer Discov. 2016;6(5):546–559. doi: 10.1158/2159-8290.CD-15-1408. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
