

NOTCH, ERK, and SHH signaling respectively control the fate determination of cortical glia and olfactory bulb interneurons
- PMID: 39999176
- PMCID: PMC11892625
- DOI: 10.1073/pnas.2416757122
NOTCH, ERK, and SHH signaling respectively control the fate determination of cortical glia and olfactory bulb interneurons
Abstract
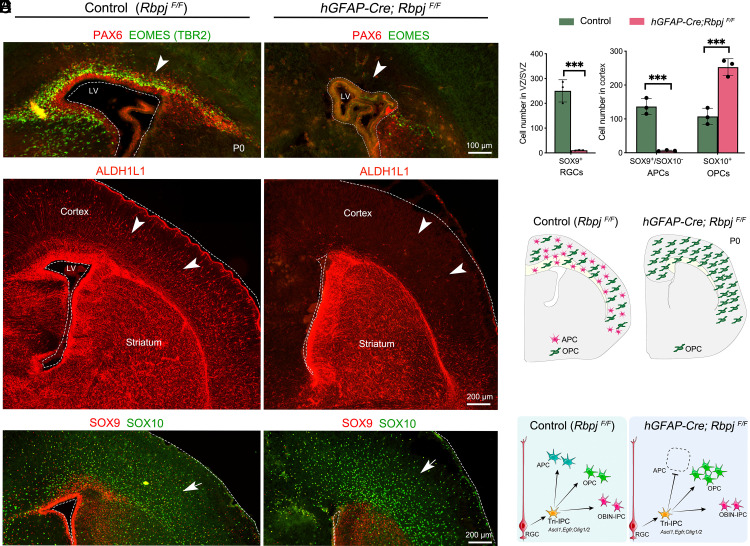
During cortical development, radial glial cells (neural stem cells) initially are neurogenic, generating intermediate progenitor cells that exclusively produce glutamatergic pyramidal neurons. Next, radial glial cells generate tripotential intermediate progenitor cells (Tri-IPCs) that give rise to cortical astrocytes and oligodendrocytes, and olfactory bulb interneurons. The molecular mechanisms underlying the transition from cortical neurogenesis to gliogenesis, and the subsequent fate determination of cortical astrocytes, oligodendrocytes, and olfactory bulb interneurons, remain unclear. Here, we report that extracellular signal-regulated kinase (ERK) signaling plays a fundamental role in promoting cortical gliogenesis and the generation of Tri-IPCs. Additionally, sonic hedgehog-smoothened-glioma-associated oncogene homolog (SHH-SMO-GLI) activator signaling has an auxiliary function to ERK during these processes. We further demonstrate that, from Tri-IPCs, NOTCH signaling is crucial for the fate determination of astrocytes, while ERK signaling plays a prominent role in oligodendrocyte fate specification, and SHH signaling is required for the fate determination of olfactory bulb interneurons. We provide evidence suggesting that this mechanism is conserved in both mice and humans. Finally, we propose a unifying principle of mammalian cortical gliogenesis.
Keywords: EGFR; ERK signaling; NOTCH signaling; SHH signaling; cortical gliogenesis.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures




References
MeSH terms
Substances
Grants and funding
- R01 NS089777/NS/NINDS NIH HHS/United States
- NSFC 31820103006 32070971 32100768 32200776 and 32200792/MOST | National Natural Science Foundation of China (NSFC)
- R01MH094589 and R01NS089777/HHS | NIH (NIH)
- R01 MH094589/MH/NIMH NIH HHS/United States
- STI2030-2021ZD0202300/Ministry of Science and Technology of the People's Republic of China (MOST)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
