

Probing aspects of extracellular vesicle associated AAV allows increased vector yield and insight into its transduction and immune-evasive properties
- PMID: 40008086
- PMCID: PMC11851286
- DOI: 10.1016/j.omtm.2025.101407
Probing aspects of extracellular vesicle associated AAV allows increased vector yield and insight into its transduction and immune-evasive properties
Abstract
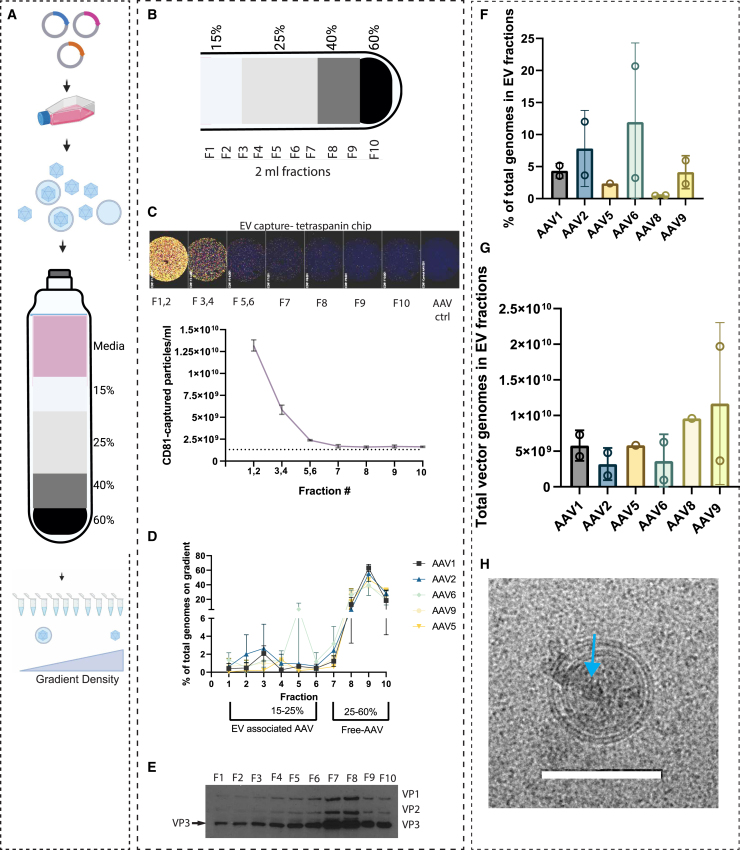
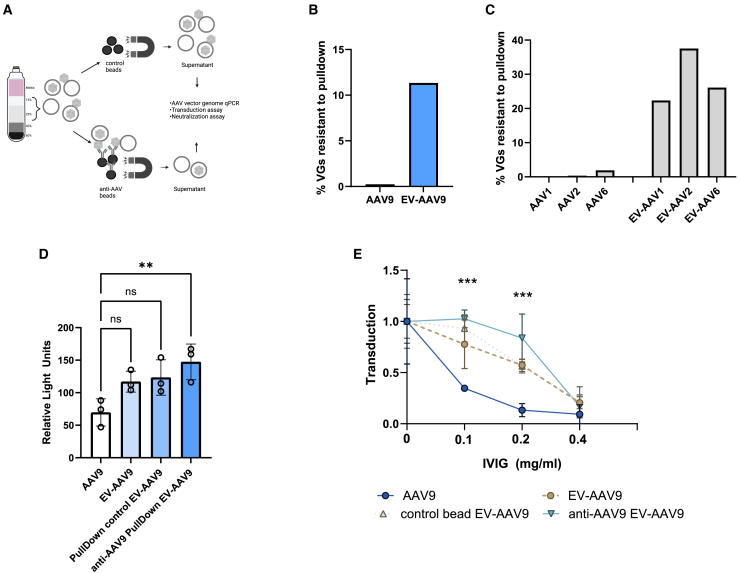
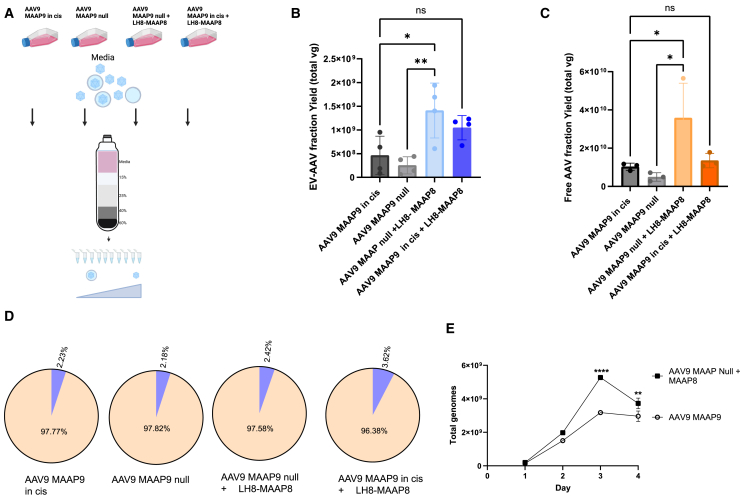
Extracellular vesicle-associated adeno-associated virus vectors (EV-AAVs) are generated during production in 293 cells. EV-AAV provides desirable gene delivery traits such as greater resistance to antibody neutralization and increased transduction of organs in vivo compared with conventional AAV. Despite these promising data, better characterization of EV-AAV is needed. We used density gradient ultracentrifugation to separate EV-AAV from free AAV to determine the yields and functional activity of EV-AAV. We found that the fraction of EV-AAV to conventional AAV in culture media from six AAV serotypes ranged from 0.5% to 12%. Next, we assessed whether intraluminal EV-AAV9 could mediate functional transduction of cells and observed that a portion of EV-AAV9 are intraluminal and mediated transduction of cultured cells in vitro and in vivo and evade antibodies compared with conventional AAV9. We tested whether trans-expression of membrane-associated accessory protein (MAAP) from AAV8 (MAAP8) or AAV9 (MAAP9) with AAV9 Cap/AAV9 MAAP null would alter yields of EV-AAV9. Trans-expression of MAAP8 or MAAP9 increased yields of EV-AAV9 compared with the cis-expression of AAV9 Cap/AAV9 MAAP. Finally, we found that the capsid was required for efficient transduction of cultured cells by EV-AAV. In sum, these data provide a foundation for the development of EV-AAV vectors.
Keywords: AAV vectors; EV-AAV; EV-associated AAV; EVs; adeno-associated virus vectors; exosomes; extracellular vesicles; gene delivery.
© 2025 The Author(s).
Conflict of interest statement
C.A.M. has financial interests in Chameleon Biosciences, Skylark Bio, and Sphere Gene Therapeutics, companies developing adeno-associated virus (AAV) vector technologies for gene therapy applications. C.A.M. performs paid consulting work for all three companies. C.A.M. has pending and issued patents surrounding EV-AAV technology, which has been licensed to biotechnology companies. C.A.M.’s interests were reviewed and are managed by Massachusetts General Hospital and Mass General Brigham in accordance with their conflict-of-interest policies.
Figures



Similar articles
-
Adeno-associated virus 9 (AAV9) viral proteins VP1, VP2, and membrane-associated accessory protein (MAAP) differentially influence in vivo transgene expression.J Virol. 2024 Nov 19;98(11):e0168124. doi: 10.1128/jvi.01681-24. Epub 2024 Oct 30. J Virol. 2024. PMID: 39475275 Free PMC article.
-
Extracellular Vesicle-Encapsulated Adeno-Associated Viruses for Therapeutic Gene Delivery to the Heart.Circulation. 2023 Aug;148(5):405-425. doi: 10.1161/CIRCULATIONAHA.122.063759. Epub 2023 Jul 6. Circulation. 2023. PMID: 37409482
-
Metabolic engineering improves transduction efficiency and downstream vector isolation by altering the lipid composition of extracellular vesicle-enclosed AAV.Metab Eng. 2025 Mar;88:40-49. doi: 10.1016/j.ymben.2024.12.003. Epub 2024 Dec 7. Metab Eng. 2025. PMID: 39653070
-
Extracellular vesicles: nature's nanoparticles for improving gene transfer with adeno-associated virus vectors.Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2018 May;10(3):e1488. doi: 10.1002/wnan.1488. Epub 2017 Aug 11. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2018. PMID: 28799250 Review.
-
Therapeutic Application of Extracellular Vesicles-Capsulated Adeno-Associated Virus Vector via nSMase2/Smpd3, Satellite, and Immune Cells in Duchenne Muscular Dystrophy.Int J Mol Sci. 2022 Jan 28;23(3):1551. doi: 10.3390/ijms23031551. Int J Mol Sci. 2022. PMID: 35163475 Free PMC article. Review.
Cited by
-
Distinguishing Protein and Gene Delivery Enables Characterization and Bioengineering of Extracellular Vesicle-Adeno-Associated Virus Vectors.bioRxiv [Preprint]. 2025 Jul 4:2025.07.02.662894. doi: 10.1101/2025.07.02.662894. bioRxiv. 2025. PMID: 40631339 Free PMC article. Preprint.
References
-
- Manno C.S., Pierce G.F., Arruda V.R., Glader B., Ragni M., Rasko J.J., Ozelo M.C., Hoots K., Blatt P., Konkle B., et al. Successful transduction of liver in hemophilia by AAV-Factor IX and limitations imposed by the host immune response. Nat. Med. 2006;12:342–347. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
