

Cryo-EM structure of the human THIK-1 K2P K+ channel reveals a lower Y gate regulated by lipids and anesthetics
- PMID: 40011745
- PMCID: PMC12263422
- DOI: 10.1038/s41594-025-01497-6
Cryo-EM structure of the human THIK-1 K2P K+ channel reveals a lower Y gate regulated by lipids and anesthetics
Abstract
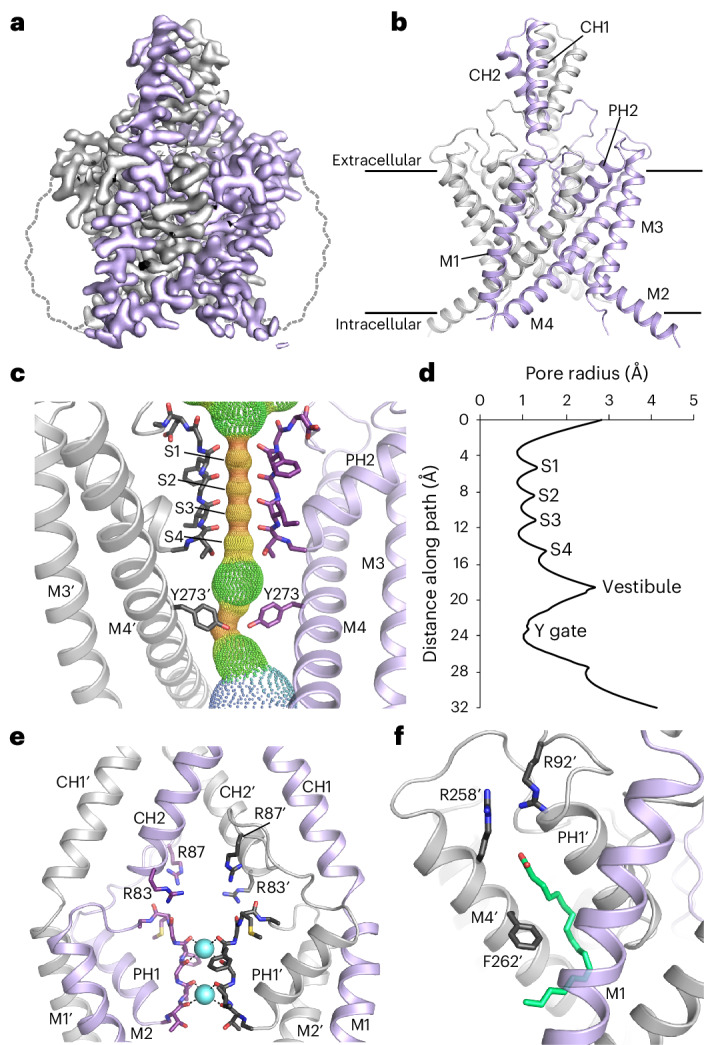
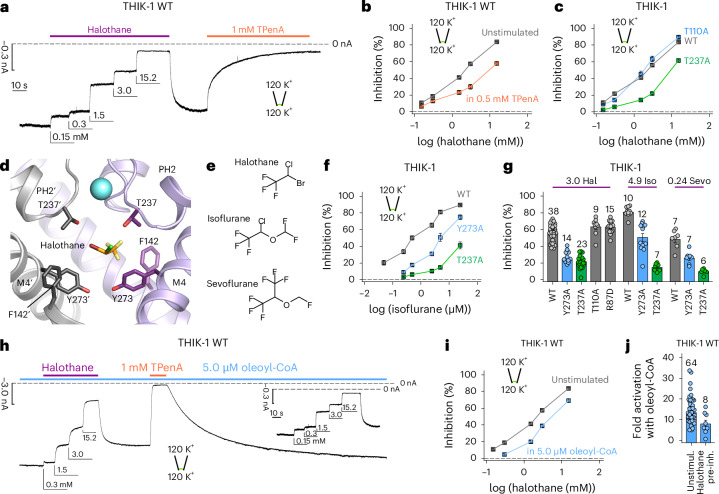
THIK-1 (KCNK13) is a halothane-inhibited and anionic-lipid-activated two-pore domain (K2P) K+ channel implicated in microglial activation and neuroinflammation, and a current target for the treatment of neurodegenerative disorders, for example Alzheimer's disease and amyothropic lateral sclerosis (ALS). However, compared to other K2P channels, little is known about the structural and functional properties of THIK-1. Here we present a 3.16-Å-resolution cryo-EM structure of human THIK-1 that reveals several distinct features, in particular, a tyrosine in M4 that contributes to a lower 'Y gate' that opens upon activation by physiologically relevant G-protein-coupled receptor and lipid signaling pathways. We demonstrate that linoleic acid bound within a modulatory pocket adjacent to the filter influences channel activity, and that halothane inhibition involves a binding site within the inner cavity, both resulting in conformational changes to the Y gate. Finally, the extracellular cap domain contains positively charged residues that line the ion exit pathway and contribute to the distinct biophysical properties of this channel. Overall, our results provide structural insights into THIK-1 function and identify distinct regulatory sites that expand its potential as a drug target for the modulation of microglial function.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: A.R. is employed by Cerevance. The other authors declare no competing interests.
Figures







References
-
- Enyedi, P. & Czirjak, G. Molecular background of leak K+ currents: two-pore domain potassium channels. Physiol. Rev.90, 559–605 (2010). - PubMed
-
- Niemeyer, M. I., Cid, L. P., Gonzalez, W. & Sepulveda, F. V. Gating, regulation, and structure in K2P K+ channels: in varietate concordia? Mol. Pharmacol.90, 309–317 (2016). - PubMed
-
- Mathie, A., Veale, E. L., Cunningham, K. P., Holden, R. G. & Wright, P. D. Two-pore domain potassium channels as drug targets: anesthesia and beyond. Annu. Rev. Pharmacol. Toxicol.61, 401–420 (2021). - PubMed
-
- Rajan, S. et al. THIK-1 and THIK-2, a novel subfamily of tandem pore domain K+ channels. J. Biol. Chem.276, 7302–7311 (2001). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
