

Glycocalyx dysregulation impairs blood-brain barrier in ageing and disease
- PMID: 40011765
- PMCID: PMC11946907
- DOI: 10.1038/s41586-025-08589-9
Glycocalyx dysregulation impairs blood-brain barrier in ageing and disease
Abstract
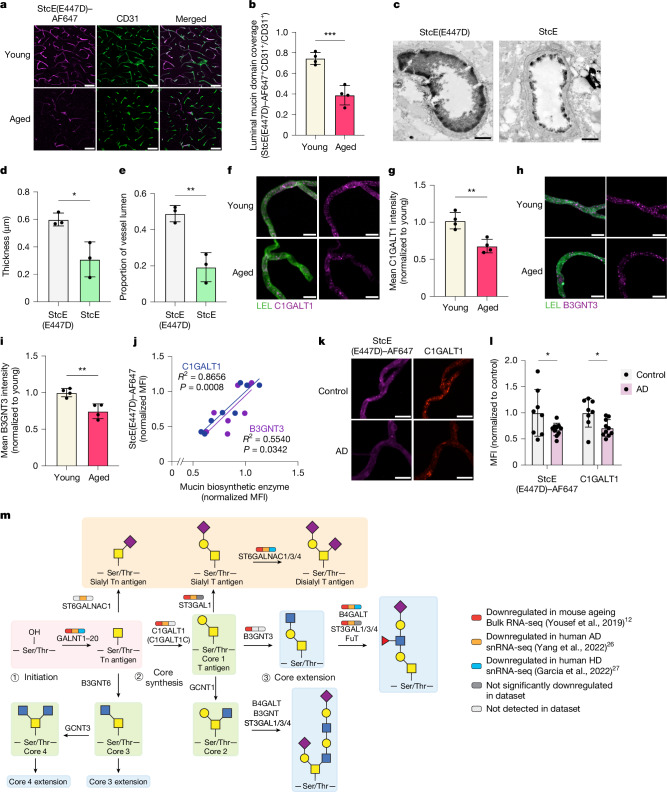
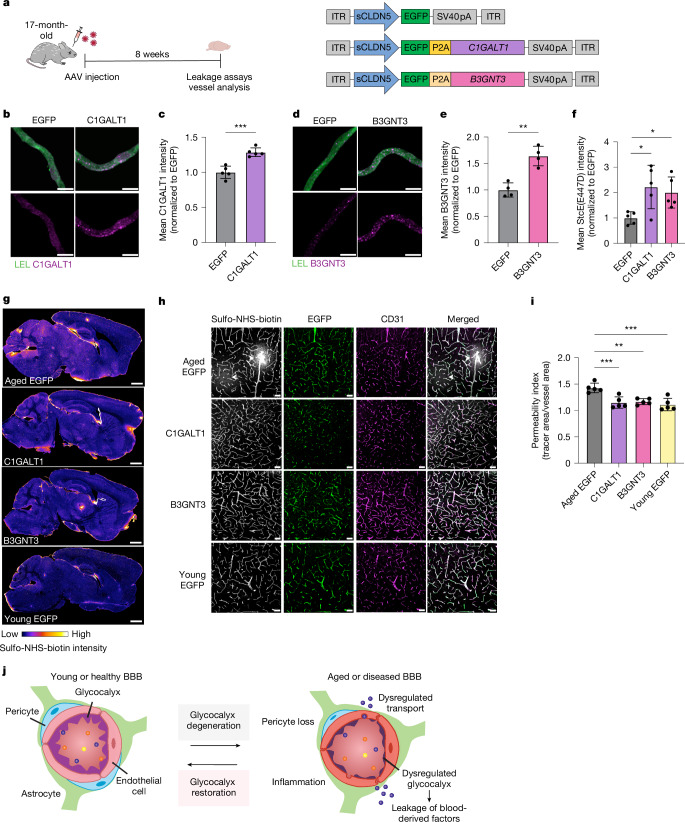
The blood-brain barrier (BBB) is highly specialized to protect the brain from harmful circulating factors in the blood and maintain brain homeostasis1,2. The brain endothelial glycocalyx layer, a carbohydrate-rich meshwork composed primarily of proteoglycans, glycoproteins and glycolipids that coats the BBB lumen, is a key structural component of the BBB3,4. This layer forms the first interface between the blood and brain vasculature, yet little is known about its composition and roles in supporting BBB function in homeostatic and diseased states. Here we find that the brain endothelial glycocalyx is highly dysregulated during ageing and neurodegenerative disease. We identify significant perturbation in an underexplored class of densely O-glycosylated proteins known as mucin-domain glycoproteins. We demonstrate that ageing- and disease-associated aberrations in brain endothelial mucin-domain glycoproteins lead to dysregulated BBB function and, in severe cases, brain haemorrhaging in mice. Finally, we demonstrate that we can improve BBB function and reduce neuroinflammation and cognitive deficits in aged mice by restoring core 1 mucin-type O-glycans to the brain endothelium using adeno-associated viruses. Cumulatively, our findings provide a detailed compositional and structural mapping of the ageing brain endothelial glycocalyx layer and reveal important consequences of ageing- and disease-associated glycocalyx dysregulation on BBB integrity and brain health.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: T.W.-C. is a co-founder and scientific advisor of Alkahest Inc., a subsidiary of Grifols SA, Qinotto Inc., and Teal Omics Inc. C.R.B. is a co-founder and scientific advisory board member of Lycia Therapeutics, Palleon Pharmaceuticals, Enable Bioscience, Redwood Biosciences (a subsidiary of Catalent), OliLux Bio, Grace Science LLC, InterVenn Biosciences, Neuravid Therapeutics, GanNA Bio, Valora Therapeutics and Firefly Biologics. S.M.S., T.W.-C. and C.R.B. are co-inventors on a patent application related to the work published in this paper. The other authors declare no competing interests.
Figures













References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
