

PPP1R13L drives cervical cancer progression by suppressing p63-mediated PTEN transcription
- PMID: 40014097
- PMCID: PMC11868476
- DOI: 10.1007/s00018-025-05598-9
PPP1R13L drives cervical cancer progression by suppressing p63-mediated PTEN transcription
Abstract
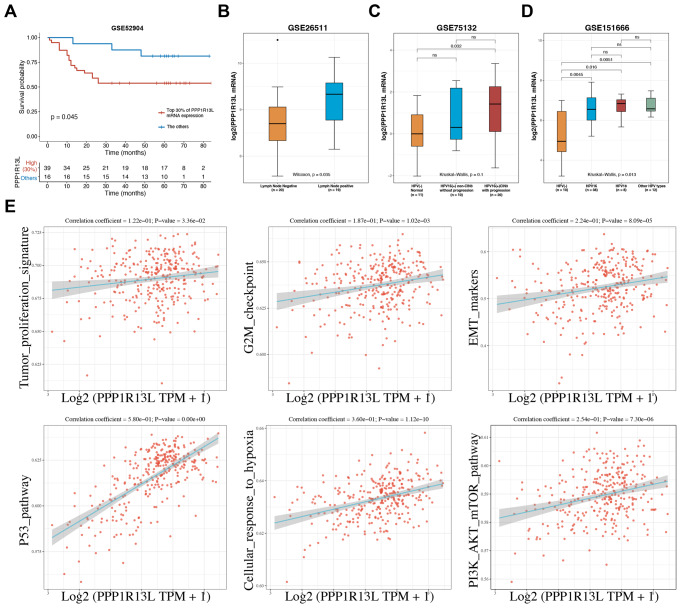
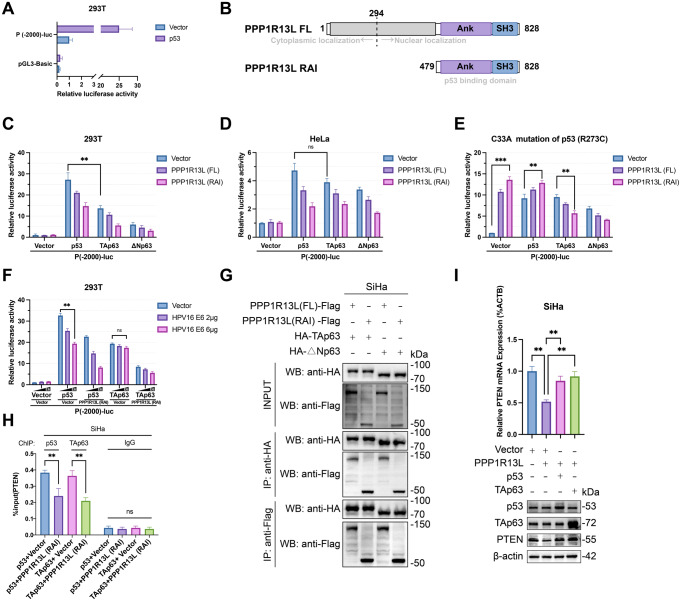
PPP1R13L is a conserved inhibitor of p53, selectively regulating a subset of p53 target genes. Previous studies have reported that PPP1R13L promotes cervical cancer progression, yet its precise mechanism remains unclear and warrants further investigation. In this study, we utilized public databases to reveal the correlation between PPP1R13L and tumor progression pathways. Subsequently, we performed functional assays both in vitro and in xenograft models to assess the impact of PPP1R13L on cervical cancer. Our results demonstrate that PPP1R13L promotes cervical cancer cell proliferation, epithelial-mesenchymal transition, cycle progression, and glycolysis via the PTEN/AKT/mTOR pathway. Mechanistically, PPP1R13L regulates the transcription of PTEN through its Ank-SH3 domain interaction with p53 family, p53 and p63. In 293T cells, p53 originally exhibits significantly higher transcriptional activity than p63. However, in cervical cancer-where E6 continuously degrades p53 and p63 is highly expressed-p63 demonstrates a transcriptional activity for PTEN that is comparable to, or even surpasses, that of p53, depending on E6 expression levels. Additionally, in C33A, an HPV-negative cervical cancer cell line, the p53 R273C mutation causes PPP1R13L to exert an opposite effect, and p63 is shown to be inhibited by PPP1R13L independently of p53. Finally, the response elements of PPP1R13L-regulated p53 family target genes were experimentally validated on p63 for the first time. This provides a sequence basis for the selective regulation of p53 family target genes by PPP1R13L. In summary, we underscore the specificity of the PPP1R13L/p63/PTEN axis in cervical cancer and propose that PPP1R13L holds potential as a therapeutic target for cervical cancer treatment.
Keywords: Cervical cancer; Glycolysis; PPP1R13L; PTEN; p63.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethical approval: This study was performed in line with the principles of the Declaration of Helsinki. Approval was granted by the Medical Ethics Committee of Zhongnan Hospital of Wuhan University (Ethics Approval No. 2020029). Consent to participate: Informed consent was obtained from all individual participants included in the study. Consent to publish: The authors affirm that human research participants provided informed consent for publication of the images in Fig. 5G. Competing interests: The authors have no relevant financial or non-financial interests to disclose. Cell lines: The human cervical cancer cell lines HeLa (CL-0101), SiHa (CL-0210), C33A (CL-0045), and the human embryonic kidney cell line 293T (CL-0005) used in this study were purchased from Procell, Wuhan, China, and were verified by STR profiling.
Figures



Similar articles
-
Tumor Suppressor LINC02487 Inhibits the Progression of Cervical Cancer in Vitro by Regulating the PTEN/Akt/mTOR Pathway.Discov Med. 2024 Aug;36(187):1732-1742. doi: 10.24976/Discov.Med.202436187.159. Discov Med. 2024. PMID: 39190388
-
LINC00861 inhibits the progression of cervical cancer cells by functioning as a ceRNA for miR‑513b‑5p and regulating the PTEN/AKT/mTOR signaling pathway.Mol Med Rep. 2021 Jan;23(1):24. doi: 10.3892/mmr.2020.11662. Epub 2020 Nov 12. Mol Med Rep. 2021. PMID: 33179755 Free PMC article.
-
PRDM4 inhibits cell proliferation and tumorigenesis by inactivating the PI3K/AKT signaling pathway through targeting of PTEN in cervical carcinoma.Oncogene. 2021 May;40(18):3318-3330. doi: 10.1038/s41388-021-01765-x. Epub 2021 Apr 12. Oncogene. 2021. PMID: 33846573 Free PMC article.
-
p63 is a suppressor of tumorigenesis and metastasis interacting with mutant p53.Cell Death Differ. 2011 Sep;18(9):1487-99. doi: 10.1038/cdd.2011.81. Epub 2011 Jul 15. Cell Death Differ. 2011. PMID: 21760596 Free PMC article. Review.
-
Maintaining epithelial stemness with p63.Sci Signal. 2015 Jul 28;8(387):re9. doi: 10.1126/scisignal.aaa1033. Sci Signal. 2015. PMID: 26221054 Review.
References
-
- Wei F, Georges D, Man I, Baussano I, Clifford GM (2024) Causal attribution of human papillomavirus genotypes to invasive cervical cancer worldwide: a systematic analysis of the global literature. Lancet 404:435–444. 10.1016/s0140-6736(24)01097-3 - PubMed
-
- Bergamaschi D, Samuels Y, O’Neil NJ, Trigiante G, Crook T, Hsieh JK, O’Connor DJ, Zhong S, Campargue I, Tomlinson ML, Kuwabara PE, Lu X (2003)iASPP oncoprotein is a key inhibitor of p53 conserved from worm to human. Nat Genet. 33,162–167. 10.1038/ng1070 - PubMed
-
- Bergamaschi D, Samuels Y, Sullivan A, Zvelebil M, Breyssens H, Bisso A, Del Sal G, Syed N, Smith P, Gasco M, Crook T, Lu X (2006) iASPP preferentially binds p53 proline-rich region and modulates apoptotic function of codon 72-polymorphic p53. Nat Genet 38:1133–1141. 10.1038/ng1879 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
