

The pathway of unconventional protein secretion involves CUPS and a modified trans-Golgi network
- PMID: 40015244
- PMCID: PMC11867701
- DOI: 10.1083/jcb.202312120
The pathway of unconventional protein secretion involves CUPS and a modified trans-Golgi network
Abstract
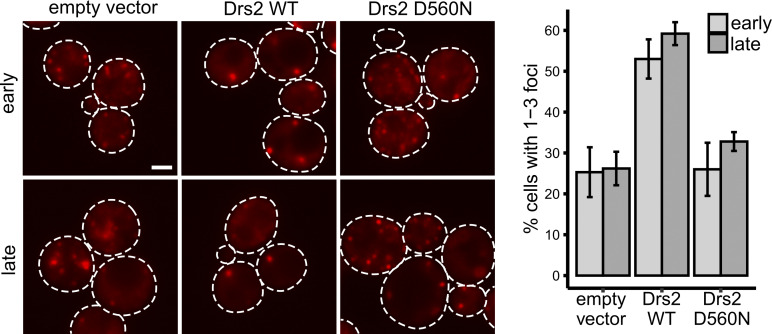
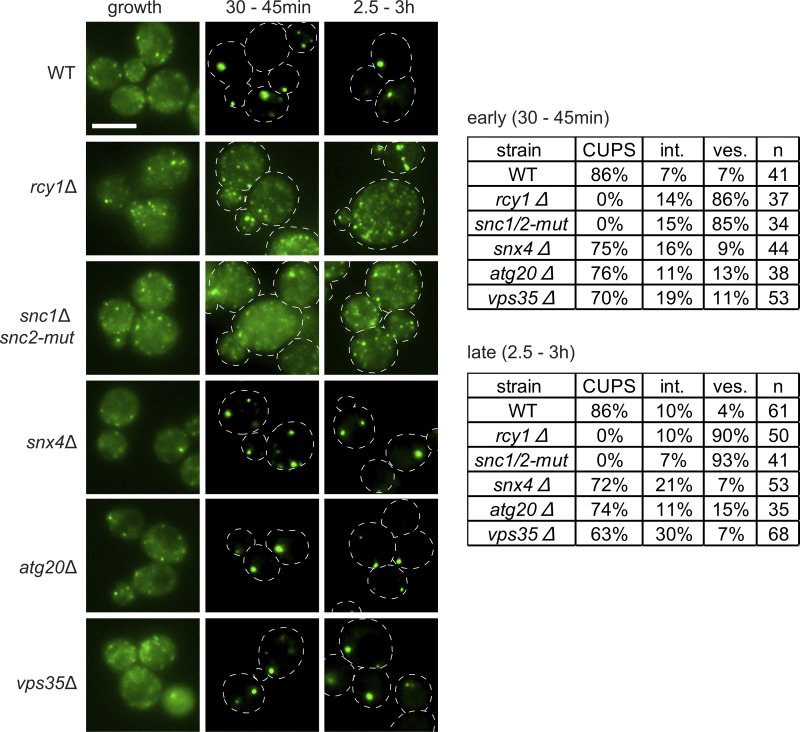
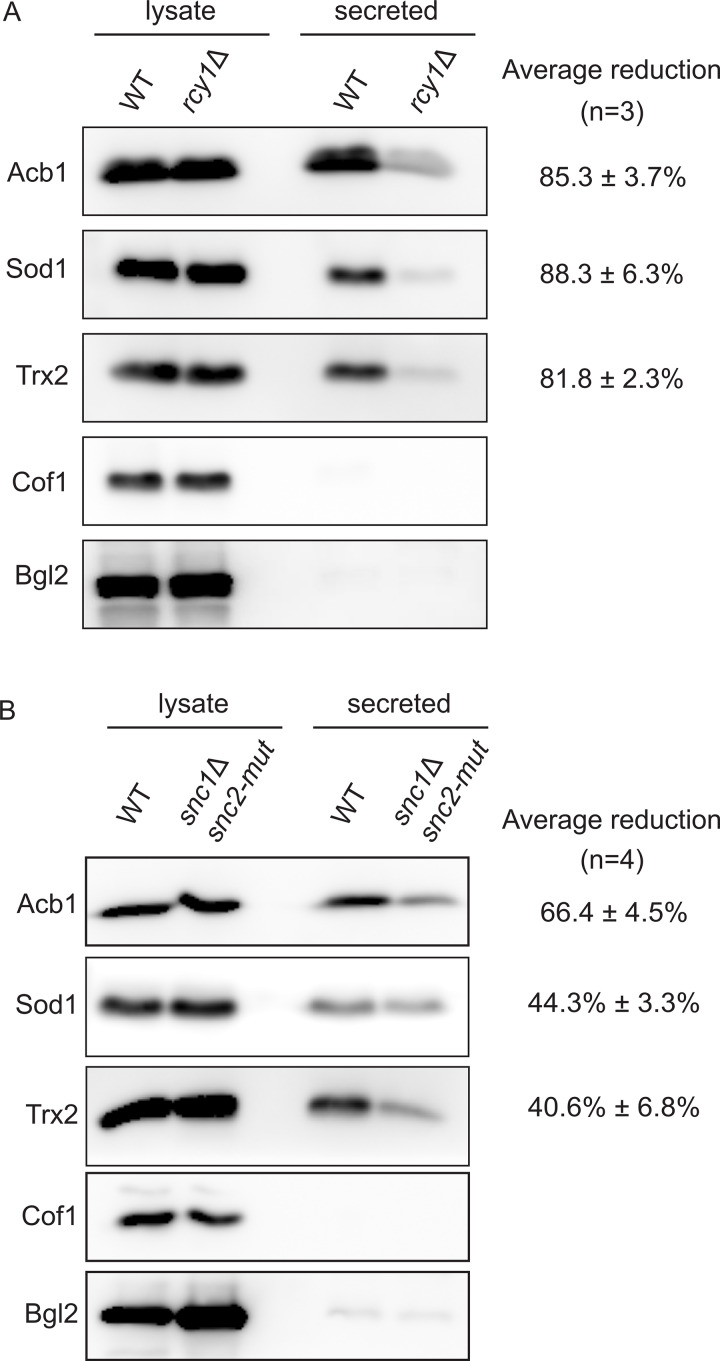
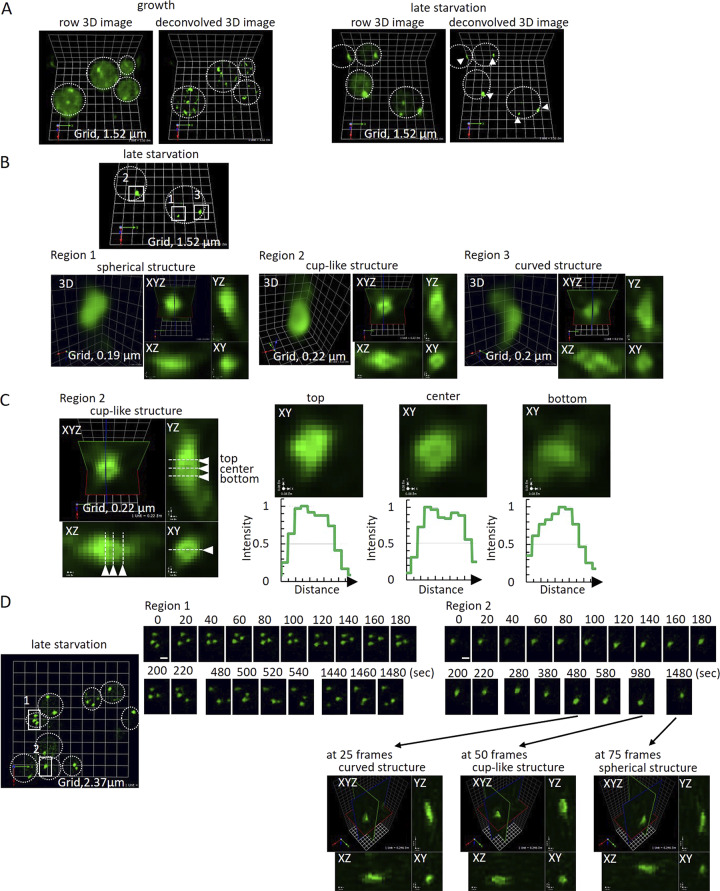
Compartment for unconventional protein secretion (CUPS), a compartment for secretion of signal sequence-lacking proteins, forms through COPI-independent extraction of membranes from early Golgi cisternae, lacks Golgi-specific glycosyltransferases, and requires phosphatidylinositol 4-phosphate (PI4P) for biogenesis, as well as phosphatidylinositol 3-phosphate for stability. Our findings demonstrate that Drs2, a PI4P effector from the trans-Golgi network (TGN), is essential for CUPS formation, specifically through its interaction with Rcy1, and Rcy1 is crucial for the unconventional secretion. Using 4D super-resolution confocal live imaging microscopy, we observed that CUPS interact with a modified TGN that contains Drs2 in addition to proteins Tlg2 and Snc2, which are necessary for membrane fusion. Notably, while CUPS remain stable, the modified TGN undergoes remodeling during the later stages of unconventional secretion. In summary, we suggest that CUPS and the modified TGN, without the function of COPII and COPI, participate in collecting and sorting unconventionally secreted proteins, reflecting the role of Golgi membranes in receiving cargo from the ER during conventional secretion.
© 2025 Curwin et al.
Conflict of interest statement
Disclosures: The authors declare no competing interests exist.
Figures








References
-
- Caielli, S., Balasubramanian P., Rodriguez-Alcazar J., Balaji U., Robinson L., Wan Z., Baisch J., Smitherman C., Walters L., Sparagana P., et al. . 2024. Type I IFN drives unconventional IL-1β secretion in lupus monocytes. Immunity. 57:2497–2513.e12. 10.1016/j.immuni.2024.09.004 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
