

Large scale investigation of GPCR molecular dynamics data uncovers allosteric sites and lateral gateways
- PMID: 40016203
- PMCID: PMC11868581
- DOI: 10.1038/s41467-025-57034-y
Large scale investigation of GPCR molecular dynamics data uncovers allosteric sites and lateral gateways
Abstract
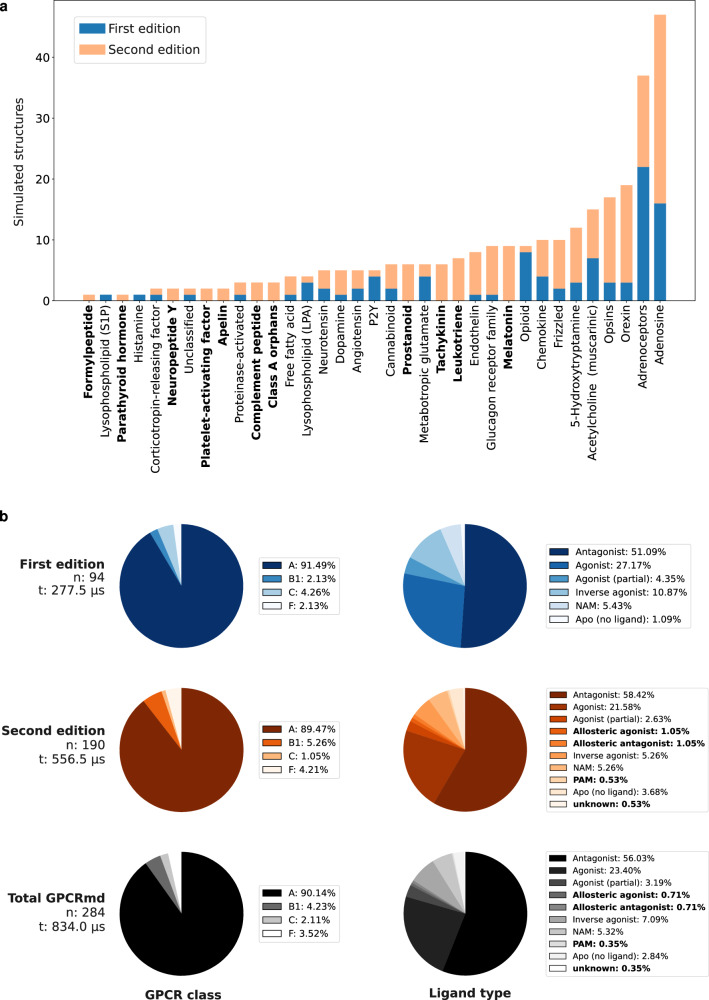
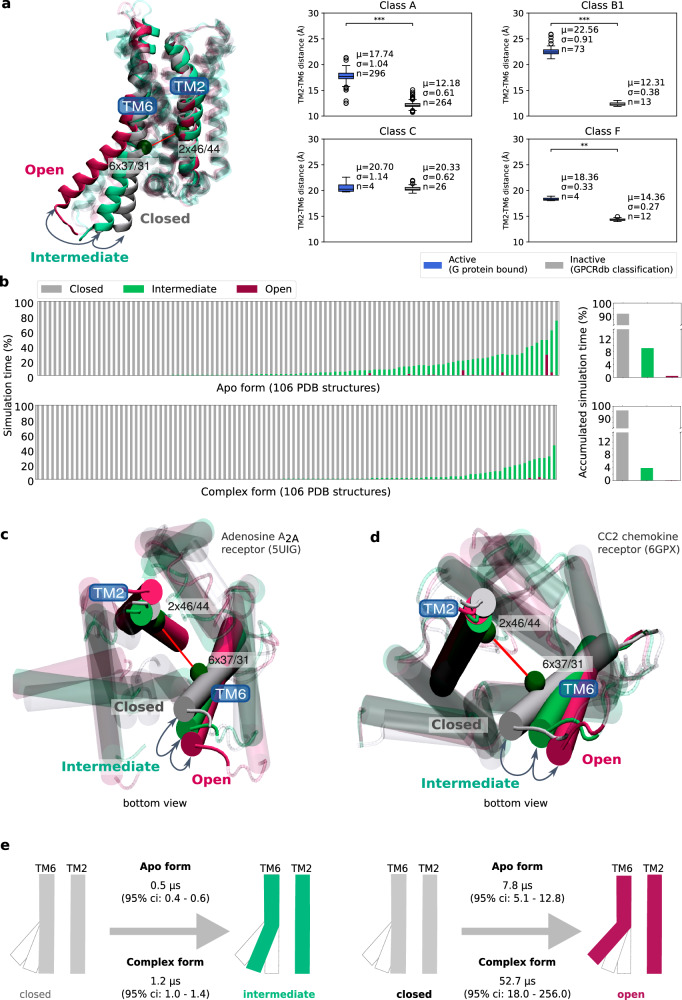
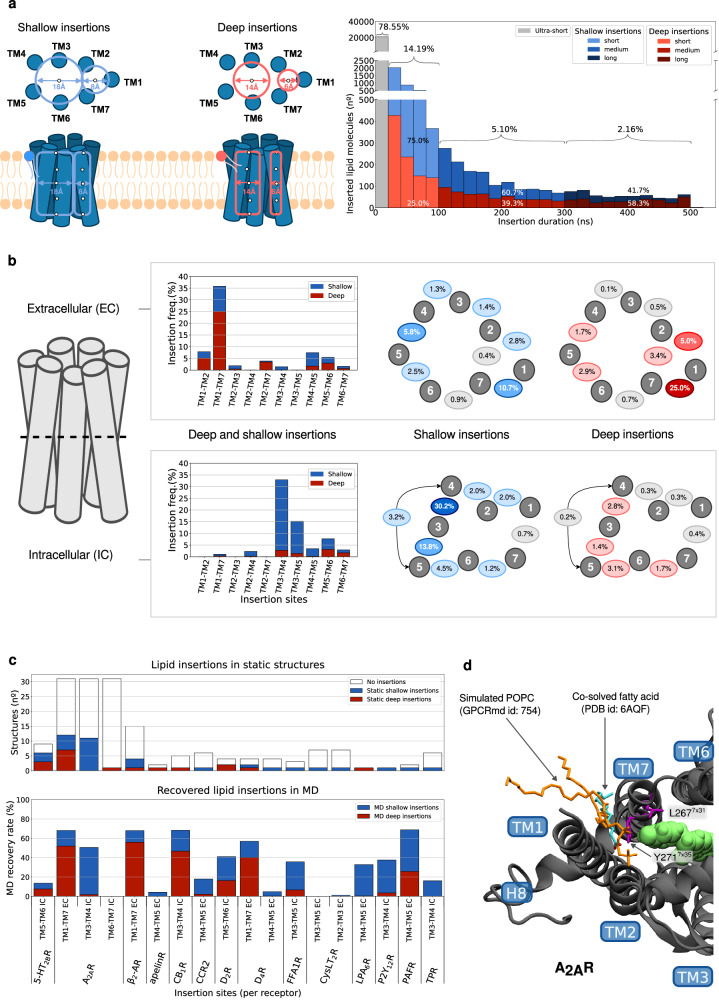
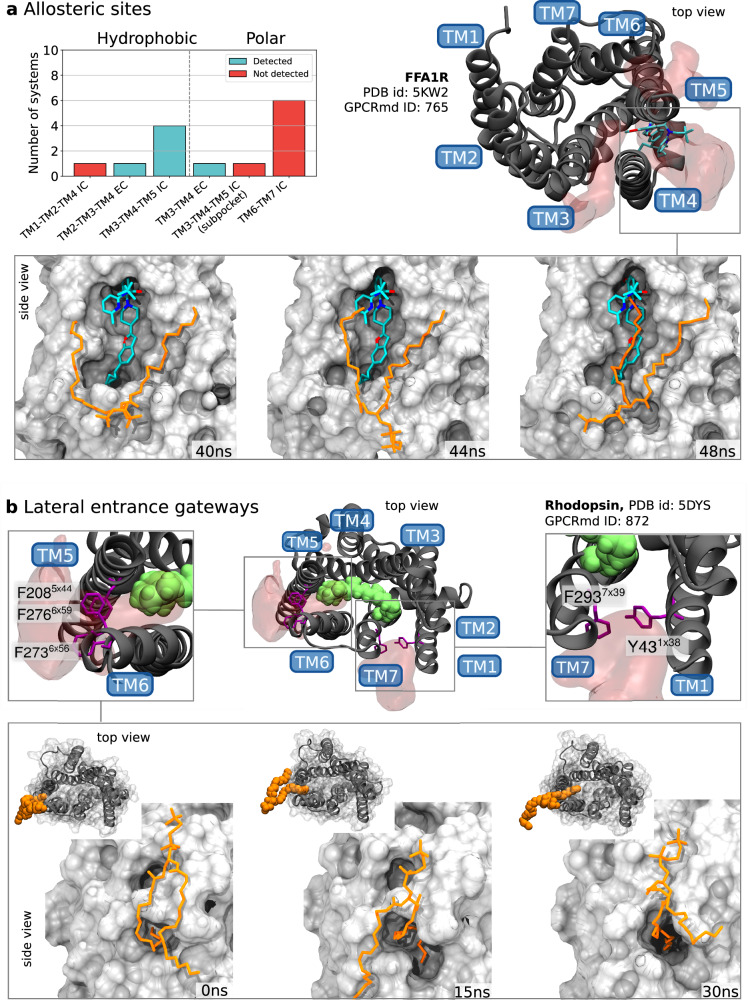
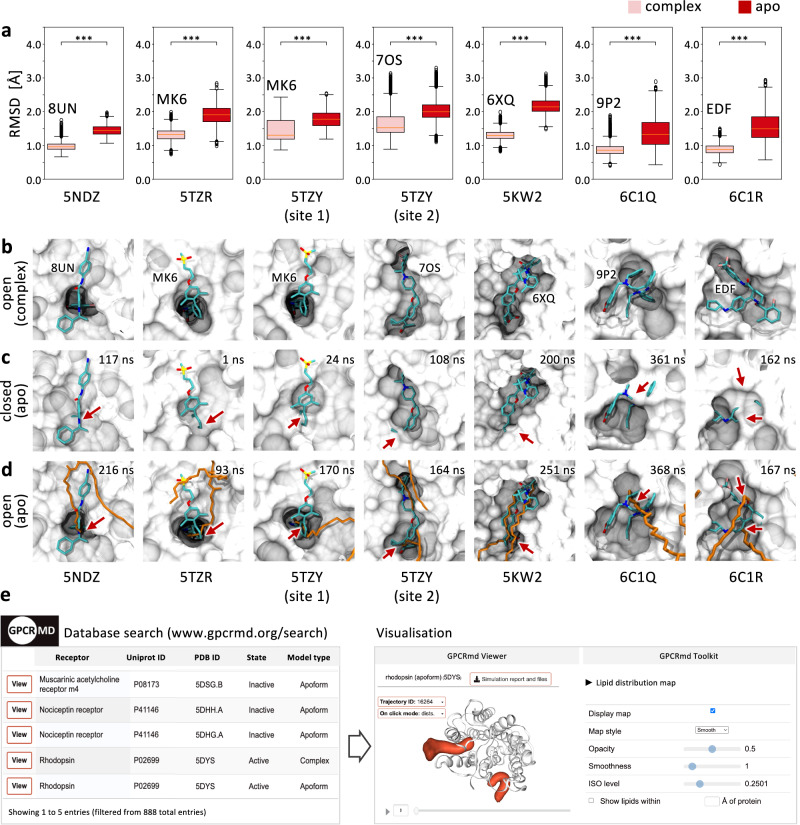
G protein-coupled receptors (GPCRs) constitute a functionally diverse protein family and are targets for a broad spectrum of pharmaceuticals. Technological progress in X-ray crystallography and cryogenic electron microscopy has enabled extensive, high-resolution structural characterisation of GPCRs in different conformational states. However, as highly dynamic events underlie GPCR signalling, a complete understanding of GPCR functionality requires insights into their conformational dynamics. Here, we present a large dataset of molecular dynamics simulations covering 60% of currently available GPCR structures. Our analysis reveals extensive local "breathing" motions of the receptor on a nano- to microsecond timescale and provides access to numerous previously unexplored receptor conformational states. Furthermore, we reveal that receptor flexibility impacts the shape of allosteric drug binding sites, which frequently adopt partially or completely closed states in the absence of a molecular modulator. We demonstrate that exploring membrane lipid dynamics and their interaction with GPCRs is an efficient approach to expose such hidden allosteric sites and even lateral ligand entrance gateways. The obtained insights and generated dataset on conformations, allosteric sites and lateral entrance gates in GPCRs allows us to better understand the functionality of these receptors and opens new therapeutic avenues for drug-targeting strategies.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
References
-
- Lopez-Balastegui, M. et al. Relevance of G protein-coupled receptor (GPCR) dynamics for receptor activation, signalling bias and allosteric modulation. Br. J. Pharmacol. 10.1111/bph.16495 (2024). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
