

Tetrandrine augments melanoma cell immunogenicity via dual inhibition of autophagic flux and proteasomal activity enhancing MHC-I presentation
- PMID: 40016522
- PMCID: PMC12205077
- DOI: 10.1038/s41401-025-01507-9
Tetrandrine augments melanoma cell immunogenicity via dual inhibition of autophagic flux and proteasomal activity enhancing MHC-I presentation
Abstract
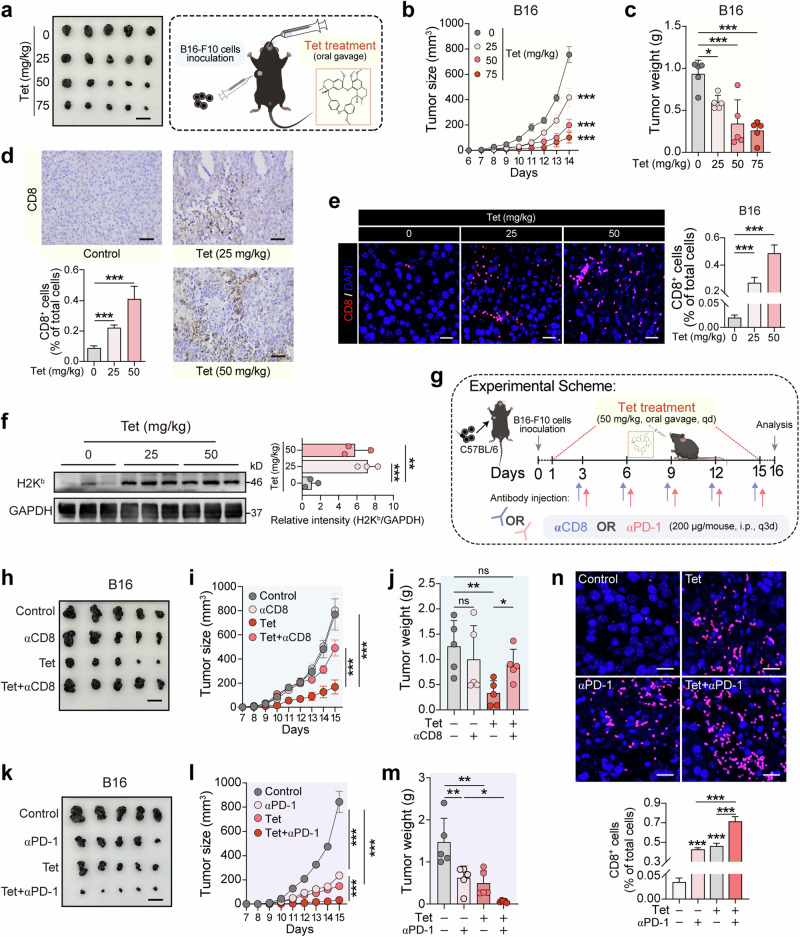
MHC-I-mediated antigen presentation is pivotal in antitumor immunity, enabling the recognition and destruction of tumor cells by CD8+ T cells. Both the proteasome and autophagy serve as essential cellular degradation mechanisms that regulate the stability and functionality of MHC-I molecules. In melanoma, modulating the pathways that affect MHC-I antigen presentation is pivotal and can profoundly influence the therapeutic outcomes of immunotherapy. Our initial effort of this study was a screening process to identify natural compounds capable of amplifying MHC-I surface expression on B16 melanoma cells. Utilizing flow cytometry with fluorescently tagged antibodies, we identified tetrandrine (Tet), a bisbenzylisoquinoline alkaloid derived from the root of Stephania tetrandra, as a potent enhancer of MHC-I-mediated antigen presentation in B16 melanoma cells. We demonstrate that tetrandrine (2.5, 5, 7.5 μM) dose-dependently upregulates both surface and total MHC-I protein levels in B16 or A375 melanoma cells by simultaneously inhibiting autophagy and proteasomal activity, two key pathways involved in MHC-I degradation. This dual inhibition stabilizes MHC-I molecules, leading to enhanced tumor antigen presentation and improved recognition by CD8+ T cells. In co-culture systems, tetrandrine treatment increased CD8+ T cell activation and cytotoxicity against melanoma cells, evidenced by elevated IFN-γ secretion and increased tumor cell apoptosis. Administration of tetrandrine (50 mg·kg-1·d-1, i.g., for 15 days) significantly suppressed melanoma growth in mouse models accompanied by increased CD8+ T cell infiltration and activation within the tumor microenvironment. Notably, tetrandrine synergized with anti-PD-1 immune checkpoint therapy, leading to enhanced tumor growth inhibition compared to either treatment alone. We revealed that tetrandrine (7.5 μM) blocked the lysosomal calcium efflux channel TPC2, disrupting lysosomal calcium homeostasis, thus impairing lysosomal acidification and proteasomal activity, thereby stabilizing MHC-I molecules and promoting antigen presentation. These results highlight tetrandrine's unique mechanism of action in enhancing MHC-I-mediated antigen presentation through dual inhibition of autophagic flux and proteasomal degradation. This study underscores tetrandrine's potential as a novel immunomodulatory agent to boost CD8+ T cell-mediated tumor cell eradication and enhance the efficacy of immune checkpoint therapies.
Keywords: MHC-I; TPC2 channel; autophagy; melanoma; proteasome; tetrandrine.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no conflict of interest.
Figures







References
-
- Mellman I, Chen DS, Powles T, Turley SJ. The cancer-immunity cycle: Indication, genotype, and immunotype. Immunity. 2023;56:2188–205. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
