

Norovirus co-opts NINJ1 for selective protein secretion
- PMID: 40020060
- PMCID: PMC11870086
- DOI: 10.1126/sciadv.adu7985
Norovirus co-opts NINJ1 for selective protein secretion
Abstract
Plasma membrane rupture by Ninjurin-1 (NINJ1) executes programmed cell death, releasing large cellular damage-associated molecular patterns (DAMPs). However, the regulation and selectivity of NINJ1-mediated DAMP release remain unexplored. Here, we uncover that murine norovirus (MNoV) strategically co-opts NINJ1 to selectively release the intracellular viral protein NS1, while NINJ1-mediated plasma membrane rupture simultaneously bulk-releases various cellular DAMPs. Host caspase-3 cleaves the precursor NS1/2, leading to NS1 secretion via an unconventional pathway. An unbiased CRISPR screen identifies NINJ1 as an essential factor for NS1 secretion. During infection, NINJ1 is recruited to the viral replication site, where it oligomerizes and forms speckled bodies, directly interacting with NS1. Subsequent mutagenesis studies identify critical amino acid residues of NS1 necessary for its interaction with NINJ1 and selective secretion. Genetic ablation or pharmaceutical inhibition of caspase-3 inhibits oral MNoV infection in mice. This study underscores the co-option of NINJ1 for controlled release of an intracellular viral protein.
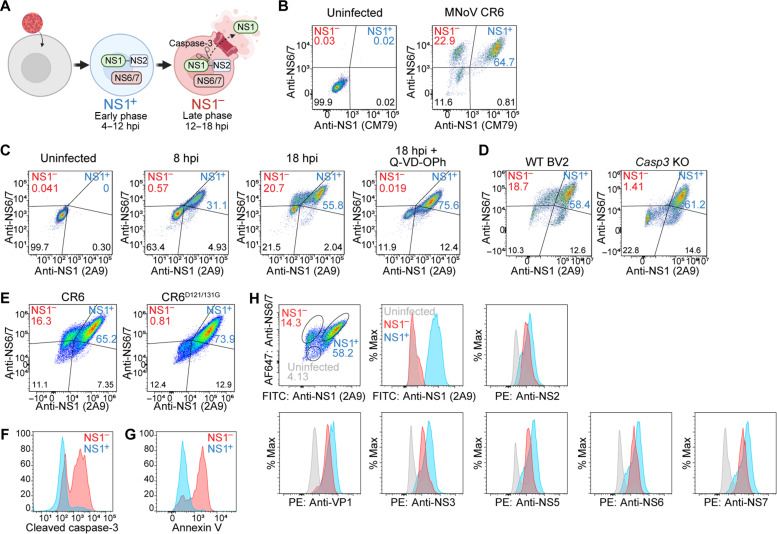
Figures





References
-
- Kayagaki N., Kornfeld O. S., Lee B. L., Stowe I. B., O’Rourke K., Li Q., Sandoval W., Yan D., Kang J., Xu M., Zhang J., Lee W. P., McKenzie B. S., Ulas G., Payandeh J., Roose-Girma M., Modrusan Z., Reja R., Sagolla M., Webster J. D., Cho V., Andrews T. D., Morris L. X., Miosge L. A., Goodnow C. C., Bertram E. M., Dixit V. M., NINJ1 mediates plasma membrane rupture during lytic cell death. Nature 591, 131–136 (2021). - PubMed
-
- Kayagaki N., Stowe I. B., Alegre K., Deshpande I., Wu S., Lin Z., Kornfeld O. S., Lee B. L., Zhang J., Liu J., Suto E., Lee W. P., Schneider K., Lin W., Seshasayee D., Bhangale T., Chalouni C., Johnson M. C., Joshi P., Mossemann J., Zhao S., Ali D., Goldenberg N. M., Sayed B. A., Steinberg B. E., Newton K., Webster J. D., Kelly R. L., Dixit V. M., Inhibiting membrane rupture with NINJ1 antibodies limits tissue injury. Nature 618, 1072–1077 (2023). - PMC - PubMed
-
- Degen M., Santos J. C., Pluhackova K., Cebrero G., Ramos S., Jankevicius G., Hartenian E., Guillerm U., Mari S. A., Kohl B., Muller D. J., Schanda P., Maier T., Perez C., Sieben C., Broz P., Hiller S., Structural basis of NINJ1-mediated plasma membrane rupture in cell death. Nature 618, 1065–1071 (2023). - PMC - PubMed
-
- Lee S., Liu H., Wilen C. B., Sychev Z. E., Desai C., Hykes B. L. Jr., Orchard R. C., McCune B. T., Kim K. W., Nice T. J., Handley S. A., Baldridge M. T., Amarasinghe G. K., Virgin H. W., A secreted viral nonstructural protein determines intestinal norovirus pathogenesis. Cell Host Microbe 25, 845–857.e5 (2019). - PMC - PubMed
MeSH terms
Substances
Grants and funding
- R01 AI183155/AI/NIAID NIH HHS/United States
- R21 AI173821/AI/NIAID NIH HHS/United States
- R01 AI148467/AI/NIAID NIH HHS/United States
- R01 HL126887/HL/NHLBI NIH HHS/United States
- R01 AI130055/AI/NIAID NIH HHS/United States
- K08 AI128043/AI/NIAID NIH HHS/United States
- U19 AI142784/AI/NIAID NIH HHS/United States
- R01 AI130020/AI/NIAID NIH HHS/United States
- R35 GM142684/GM/NIGMS NIH HHS/United States
- P20 GM109035/GM/NIGMS NIH HHS/United States
- P20 GM121344/GM/NIGMS NIH HHS/United States
- K08 AI144033/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
