

Competition for the nascent leading strand shapes the requirements for PCNA loading in the replisome
- PMID: 40021844
- PMCID: PMC12000384
- DOI: 10.1038/s44318-025-00386-4
Competition for the nascent leading strand shapes the requirements for PCNA loading in the replisome
Abstract
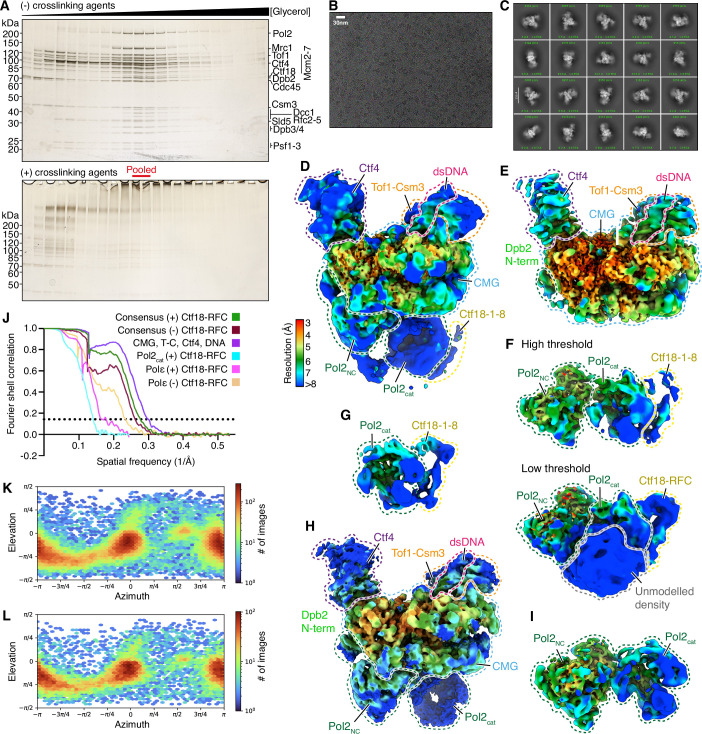
During DNA replication, the DNA polymerases Pol δ and Pol ε utilise the ring-shaped sliding clamp PCNA to enhance their processivity. PCNA loading onto DNA is accomplished by the clamp loaders RFC and Ctf18-RFC, which function primarily on the lagging and the leading strand, respectively. RFC activity is essential for lagging-strand replication by Pol δ, but it is unclear why Ctf18-RFC is required for leading-strand PCNA loading and why RFC cannot fulfil this function. Here, we show that RFC cannot load PCNA once Pol ε has been incorporated into the budding yeast replisome and commenced leading-strand synthesis, and this state is maintained during replisome progression. By contrast, we find that Ctf18-RFC is uniquely equipped to load PCNA onto the leading strand and show that this activity requires a direct interaction between Ctf18 and the CMG (Cdc45-MCM-GINS) helicase. Our work uncovers a mechanistic basis for why replisomes require a dedicated leading-strand clamp loader.
Keywords: CMG Helicase; Clamp Loader; Ctf18-RFC; PCNA Loading; Replisome.
© 2025. The Author(s).
Conflict of interest statement
Disclosure and competing interests statement. The authors declare no competing interests.
Figures











References
-
- Bambara RA, Murante RS, Henricksen LA (1997) Enzymes and reactions at the eukaryotic DNA replication fork. J Biol Chem 272:4647–4650 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
