

Glucose-dependent insulinotropic polypeptide (GIP)
- PMID: 40024571
- PMCID: PMC11931254
- DOI: 10.1016/j.molmet.2025.102118
Glucose-dependent insulinotropic polypeptide (GIP)
Abstract
Background: Glucose-dependent insulinotropic polypeptide (GIP) was the first incretin identified and plays an essential role in the maintenance of glucose tolerance in healthy humans. Until recently GIP had not been developed as a therapeutic and thus has been overshadowed by the other incretin, glucagon-like peptide 1 (GLP-1), which is the basis for several successful drugs to treat diabetes and obesity. However, there has been a rekindling of interest in GIP biology in recent years, in great part due to pharmacology demonstrating that both GIPR agonism and antagonism may be beneficial in treating obesity and diabetes. This apparent paradox has reinvigorated the field, led to new lines of investigation, and deeper understanding of GIP.
Scope of review: In this review, we provide a detailed overview on the multifaceted nature of GIP biology and discuss the therapeutic implications of GIPR signal modification on various diseases.
Major conclusions: Following its classification as an incretin hormone, GIP has emerged as a pleiotropic hormone with a variety of metabolic effects outside the endocrine pancreas. The numerous beneficial effects of GIPR signal modification render the peptide an interesting candidate for the development of pharmacotherapies to treat obesity, diabetes, drug-induced nausea and both bone and neurodegenerative disorders.
Keywords: Diabetes; GIP; GLP-1; Incretin; Insulin; Obesity.
Copyright © 2025 The Author(s). Published by Elsevier GmbH.. All rights reserved.
Conflict of interest statement
Declaration of competing interest DJD has served as a consultant or speaker within the past 12 months to Amgen, AstraZeneca, Boehringer Ingelheim, Kallyope and Novo Nordisk Inc. Neither DJD or his family members hold issued stock directly or indirectly in any of these companies. DJD holds non-exercised options in Kallyope. S.A.M, J.D.D., B.F., and P.J.K. are shareholders and former employees of Novo Nordisk. RJS has received research support from Novo Nordisk, Fractyl, Astra Zeneca, Congruence Therapeutics, Eli Lilly, Bullfrog AI, Glycsend Therapeutics and Amgen. RJS has served as a paid consultant for Novo Nordisk, Eli Lilly, CinRx, Fractyl, Structure Therapeutics, Crinetics, Amgen and Congruence Therapeutics. RJS has equity in Bullfrog AI and Rewind. MMR and JJH are co-founders and shareholders of Antag Therapeutics and Bainan Biotech. MB received honoraria as a consultant and speaker from Amgen, AstraZeneca, Bayer, Boehringer-Ingelheim, Daiichi-Sankyo, Lilly, Novo Nordisk, Novartis, Pfizer and Sanofi. DW is shareholder and on the scientific advisory board of Septerna Inc. and a co-founder and shareholder of Dacra therapeutics. MRH receives research SRA fundining from Boehringer Ingelheim, Eli Lilly & Co., Pfizer, Gila Therapeutics, and Novo Nordisk. MRH is a named inventor of patents pursuant to work that is owned by Syracuse University and the University of Pennsylvania. MRH is a founding scientists and shareholder of Coronation Bio. Inc. FMG and FR have received research support from Eli Lilly and AstraZeneca. FKK has served on scientific advisory panels, been part of speaker's bureaus for, served as a consultant to, owns stocks in and/or received research support from 89bio, Amgen, AstraZeneca, Boehringer Ingelheim, Carmot Therapeutics, Eli Lilly, Gubra, MedImmune, MSD/Merck, Norgine, Novo Nordisk, Sanofi, ShouTi, SNIPR Biome, Zealand Pharma and Zucara. FKK is a co-founder of and minority shareholder in Antag Therapeutics. FKK is currently employed by Novo Nordisk; the present work was done independent of Novo Nordisk. CH is a named inventor on patents that cover GIP or dual GLP-1/GIP receptor agonists as treatments for AD/PD. He is the CSO of Kariya Pharmaceutics Ltd. JJH appears on advisory boards for Novo Nordisk. SDP has served as president of EASD/European Foundation for the Study of Diabetes (EFSD) (2020–2022) and is current president of Fondazione Menarini; has received research grants to the institution from AstraZeneca and Boehringer Ingelheim; has served as advisor for Abbott, Amarin Corporation, Amplitude, Applied Therapeutics, AstraZeneca, Biomea Fusion, Eli Lilly & Co., EvaPharma, Menarini International, Novo Nordisk, Sanofi, and Sun Pharmaceuticals; and has received fees for speaking from AstraZeneca, Boehringer Ingelheim, Eli Lilly & Co., Laboratori Guidotti, Menarini International, Merck Sharpe & Dohme, and Novo Nordisk. PRF has served as consultant for Amgen, Ipsen, Novo Nordisk, Sanofi, Zealand and is co-founder and shareholder of Dia Beta Labs. MHT is a member of the scientific advisory board of ERX Pharmaceuticals, Cambridge, Mass. He was a member of the Research Cluster Advisory Panel (ReCAP) of the Novo Nordisk Foundation between 2017 and 2019. He attended a scientific advisory board meeting of the Novo Nordisk Foundation Center for Basic Metabolic Research, University of Copenhagen, in 2016. He received funding for his research projects by Novo Nordisk (2016–2020) and Sanofi-Aventis (2012–2019). He was a consultant for Bionorica SE (2013–2017), Menarini Ricerche S.p.A. (2016), and Bayer Pharma AG Berlin (2016). As former Director of the Helmholtz Diabetes Center and the Institute for Diabetes and Obesity at Helmholtz Zentrum München (2011–2018), and since 2018, as CEO of Helmholtz Zentrum München, he has been responsible for collaborations with a multitude of companies and institutions, worldwide. In this capacity, he discussed potential projects with and has signed/signs contracts for his institute(s) and for the staff for research funding and/or collaborations with industry and academia, worldwide, including but not limited to pharmaceutical corporations like Boehringer Ingelheim, Eli Lilly, Novo Nordisk, Medigene, Arbormed, BioSyngen, and others. In this role, he was/is further responsible for commercial technology transfer activities of his institute(s), including diabetes related patent portfolios of Helmholtz Zentrum München as, e.g., WO/2016/188932 A2 or WO/2017/194499 A1. MHT confirms that to the best of his knowledge none of the above funding sources were involved in the preparation of this paper. MAN has been member on advisory boards or has consulted with Boehringer Ingelheim, Eli Lilly & Co., Medtronic, Merck, Sharp & Dohme, NovoNordisk, Pfizer, Regor, Sun Pharma, and Structure Therapeutics (ShouTi, Gasherbrum). He has received grant support from Merck, Sharp & Dohme. He has also served on the speakers' bureau of Eli Lilly & Co., Merck, Sharp & Dohme, Medscape, Medical Learning Institute, and NovoNordisk. MAN has been member on advisory boards or has consulted with Boehringer Ingelheim, Eli Lilly & Co., Medtronic, Merck, Sharp & Dohme, NovoNordisk, Pfizer, Regor, Sun Pharma, and Structure Therapeutics (ShouTi, Gasherbrum). He has received grant support from Merck, Sharp & Dohme. He has also served on the speakers' bureau of Eli Lilly & Co., Merck, Sharp & Dohme, Medscape, Medical Learning Institute, and NovoNordisk. MAN has been member on advisory boards or has consulted with Boehringer Ingelheim, Eli Lilly & Co., Medtronic, Merck, Sharp & Dohme, NovoNordisk, Pfizer, Regor, Sun Pharma, and Structure Therapeutics (ShouTi, Gasherbrum). He has received grant support from Merck, Sharp & Dohme. He has also served on the speakers' bureau of Eli Lilly & Co., Merck, Sharp & Dohme, Medscape, Medical Learning Institute, and NovoNordisk. TDM receives funding from Novo Nordisk and has received speaking fees from Novo Nordisk, Eli Lilly, Boehringer Ingelheim, Merck, AstraZeneca, Mercodia and Berlin Chemie AG. TDM further holds stocks from Novo Nordisk and Eli Lilly. BA has served as a speaker within the past 12 months to Mankind and Novartis and is a shareholder of Astra Zeneca, Eli Lilly and Novo Nordisk AS. R.J.S., M.P.C., K.W.S., B.F. and R.E.G. are employees of Eli Lilly and Company and may own company stock.
Figures

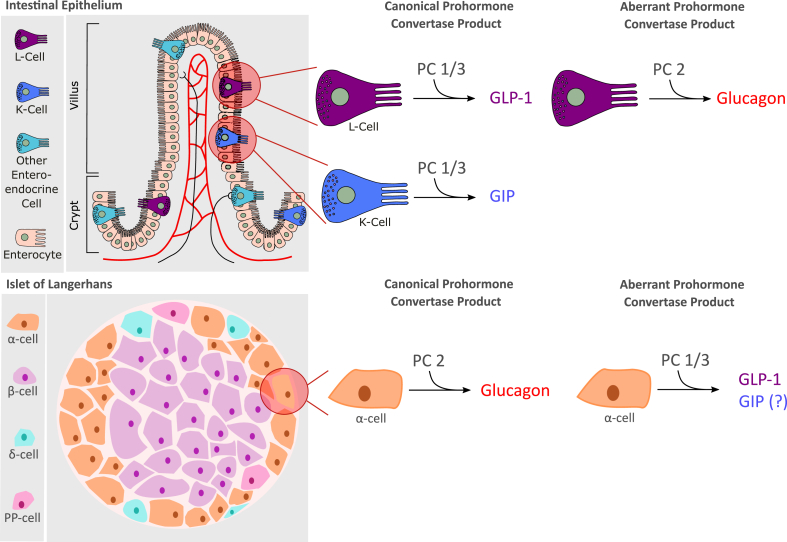

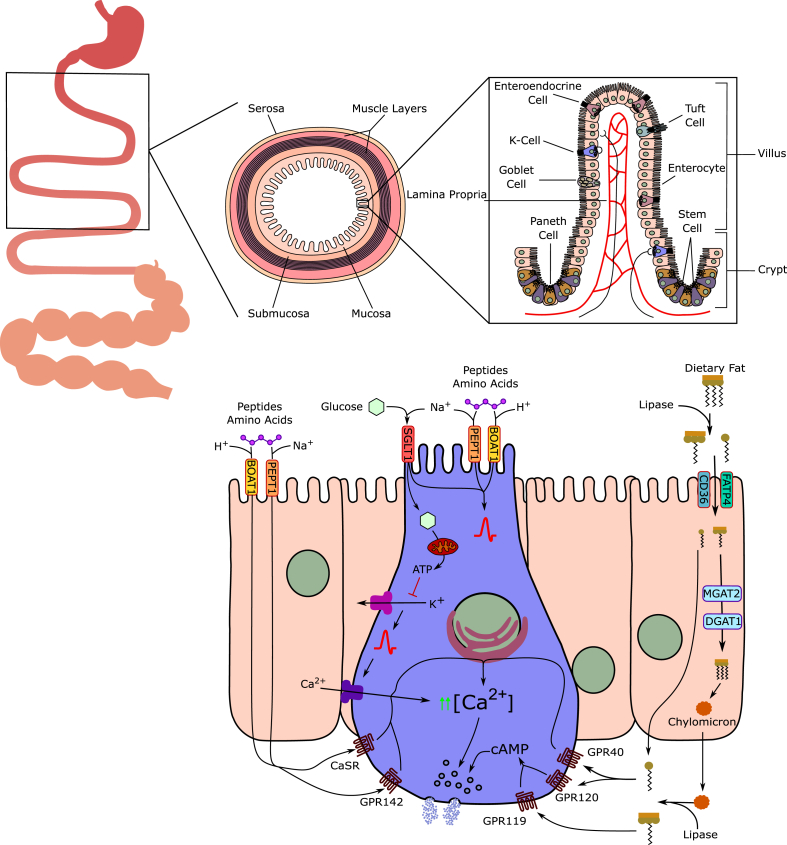
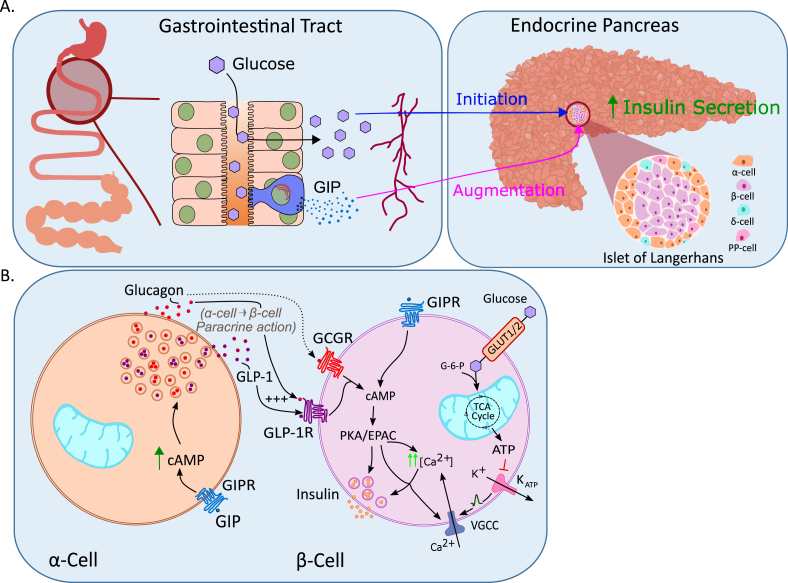
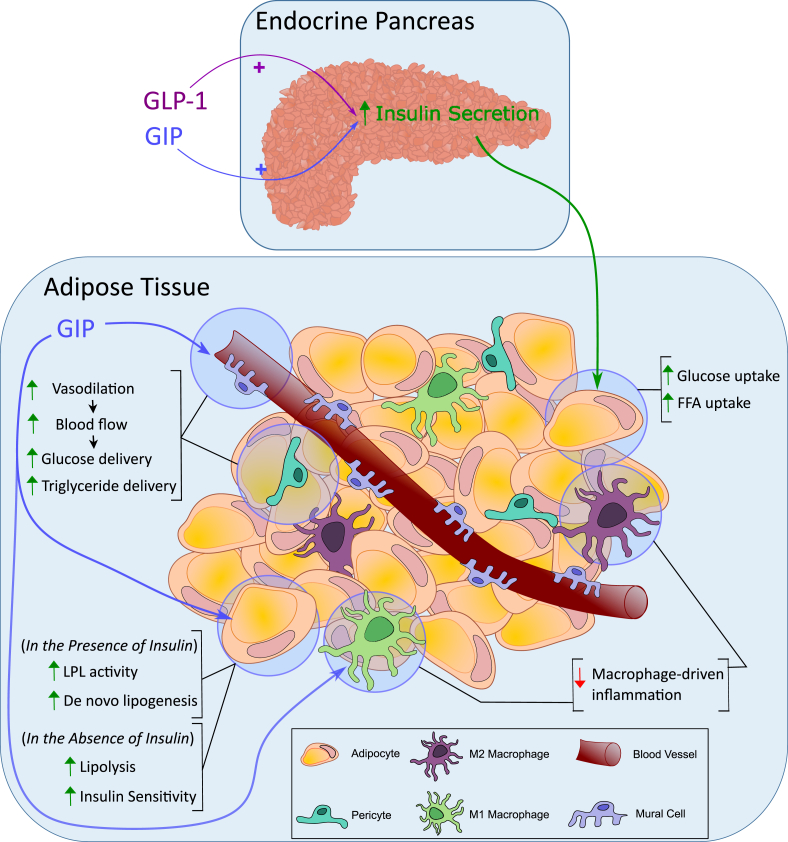


References
-
- Pawlow I.P. Walther. Verlag: Wiesbaden, JF Bergmann; 1898. Die Arbeit der Verdauungsdrüsen. Translated from Russian by A.
-
- Popielski L. Ueber das peripherische rēectorische Nervenzentrum des Pankreas. Pflügers Arch Ges Physiol. 1901;86:215–245.
-
- Wertheimer E., Lepage L. Secretion pancreatique et atropine. Comp rend Soc de biol. 1901;759
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
