

A "Dock-Work" Orange: A Dual-Receptor Biochemical Theory on the Deterrence Induced by Citrusy Aroma on Elephant Traffic Central to a Conservation Effort
- PMID: 40026377
- PMCID: PMC11869256
- DOI: 10.1177/11779322251315922
A "Dock-Work" Orange: A Dual-Receptor Biochemical Theory on the Deterrence Induced by Citrusy Aroma on Elephant Traffic Central to a Conservation Effort
Abstract
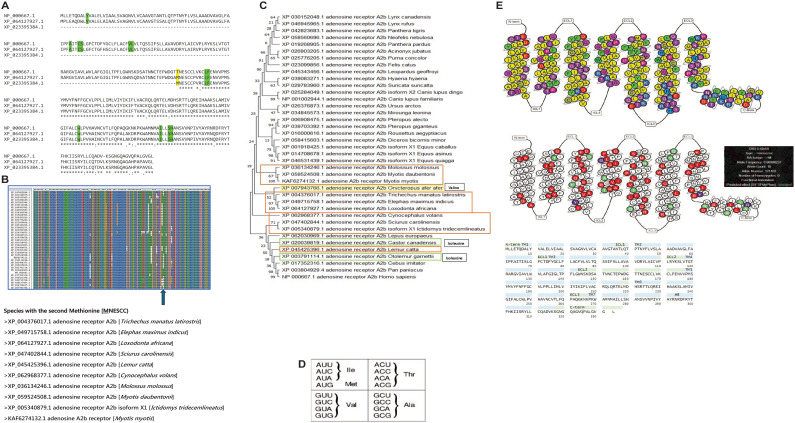
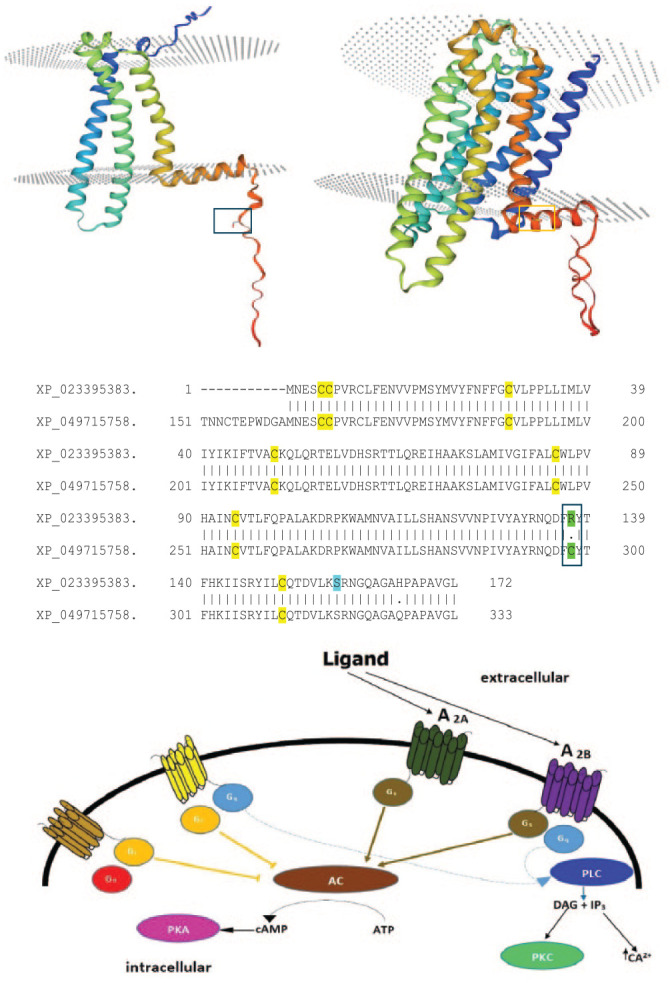
Conservation of elephants requires physical, chemical, and biological approaches to ensure the protection of these gargantuan pachyderms. One such approach is using orange plants (as biofencing) for the repellence of elephants, which precludes catastrophic events related to the encroachment of elephants into human habitats. Elephants have sensitive olfactory discrimination of plant volatile compounds for foraging and other behavior using G-protein-coupled receptors (GPCRs). However, 2 such receptors are the A2A and A2B receptors mediating olfaction elicited by a host of ligands, including limonene, the main volatile compound in citrus plants, which is hypothesized to be the chief repelling agent. Bioinformatics at the protein and mRNA levels (BLAST/Multiple Sequence Alignments) were employed to explore the multiple expression products of A2B receptors, namely full-length and truncated proteins produced by isoform mRNAs translated from multiple methionines, while the comparison of the limonene-binding pockets of human and elephant A2B receptors and prediction servers [Netphos 3.1; Protter] was used to focus, respectively, on the contacts limonene binding entails and the post-translational modifications that are involved in cell signaling. Finally, the link between limonene and antifeedant activity was explored by considering limonene content on trees that are preferentially foraged or avoided as part of the feeding behavior by elephants. The African bush elephant (Loxodonta africana) possesses a full-length A2A receptor but unlike most mammals, expresses a highly truncated A2B receptor isoform possessing only transmembrane helices 5, 6, and 7. Truncation may lead to higher traffic and expression of the A2B receptor in olfactory interfaces/pathways and aid stronger activation. In addition, all residues in the putative limonene-binding cleft are perfectly conserved between the human and African bush elephant A2B receptors, both full length and truncated. Shallow activation sites require micromolar affinity and fewer side-chain interactions, which is speculated to be the case for the truncated A2B receptor. An N-terminal extremity N-glycosylation motif is indicative of membrane localization of the truncated A2B receptor following accurate folding. A combination of truncation, indels, substitutions, and transcript isoforms are the attributed roles in the evolution of the L. africana A2B receptor, out of which limonene receptivity may be the key. It is also inferred how limonene may act as a dietary repellent/antifeedant to a generalist herbivore, with the documented limonene content being absent in some dietary favorites including the iconic Sclerocarya birrea.
Keywords: A2A receptors; A2B receptors; citrus; conservation; elephants; limonene.
© The Author(s) 2025.
Conflict of interest statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Figures








Similar articles
-
The deterrent effects of individual monoterpene odours on the dietary decisions of African elephants.Anim Cogn. 2023 Jun;26(3):1049-1063. doi: 10.1007/s10071-023-01755-4. Epub 2023 Feb 17. Anim Cogn. 2023. PMID: 36800131 Free PMC article.
-
Testing the Effectiveness of the "Smelly" Elephant Repellent in Controlled Experiments in Semi-Captive Asian and African Savanna Elephants.Animals (Basel). 2023 Oct 26;13(21):3334. doi: 10.3390/ani13213334. Animals (Basel). 2023. PMID: 37958089 Free PMC article.
-
Myth, marula, and elephant: an assessment of voluntary ethanol intoxication of the African elephant (Loxodonta africana) following feeding on the fruit of the marula tree (Sclerocarya birrea).Physiol Biochem Zool. 2006 Mar-Apr;79(2):363-9. doi: 10.1086/499983. Epub 2006 Feb 6. Physiol Biochem Zool. 2006. PMID: 16555195
-
Acknowledging the Relevance of Elephant Sensory Perception to Human-Elephant Conflict Mitigation.Animals (Basel). 2022 Apr 14;12(8):1018. doi: 10.3390/ani12081018. Animals (Basel). 2022. PMID: 35454264 Free PMC article. Review.
-
Do elephants really never forget? What we know about elephant memory and a call for further investigation.Learn Behav. 2025 Mar;53(1):44-64. doi: 10.3758/s13420-024-00655-y. Epub 2024 Oct 22. Learn Behav. 2025. PMID: 39438402 Review.
References
LinkOut - more resources
Full Text Sources
Research Materials
