

This is a preprint.
Coupled equilibria of dimerization and lipid binding modulate SARS Cov 2 Orf9b interactions and interferon response
- PMID: 40027672
- PMCID: PMC11870501
- DOI: 10.1101/2025.02.16.638509
Coupled equilibria of dimerization and lipid binding modulate SARS Cov 2 Orf9b interactions and interferon response
Update in
-
Coupled equilibria of dimerization and lipid binding modulate SARS Cov 2 Orf9b interactions and interferon response.Elife. 2025 Sep 17;14:RP106484. doi: 10.7554/eLife.106484. Elife. 2025. PMID: 40960494 Free PMC article.
Abstract
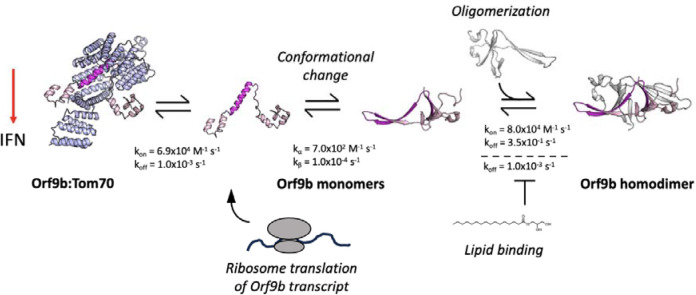
Open Reading Frame 9b (Orf9b), an accessory protein of SARS-CoV and -2, is involved in innate immune suppression through its binding to the mitochondrial receptor Translocase of Outer Membrane 70 (Tom70). Previous structural studies of Orf9b in isolation revealed a β-sheet-rich homodimer, however, structures of Orf9b in complex with Tom70 revealed a monomeric helical fold. Here, we developed a biophysical model that quantifies how Orf9b switches between these conformations and binds to Tom70, a requirement for suppressing the type 1 interferon response. We used this model to characterize the effect of lipid binding and mutations in variants of concern to the Orf9b:Tom70 equilibrium. We found that the binding of a lipid to the Orf9b homodimer biases the Orf9b monomer:dimer equilibrium towards the dimer by reducing the dimer dissociation rate ~100-fold. We also found that mutations in variants of concern can alter different microscopic rate constants without significantly affecting binding to Tom70. Together our results highlight how perturbations to different steps in these coupled equilibria can affect the apparent affinity of Orf9b to Tom70, with potential downstream implications for interferon signaling in coronavirus infection.
Conflict of interest statement
Competing Interest Statement The Krogan Laboratory has received research support from Vir Biotechnology, F. Hoffmann-La Roche, and Rezo Therapeutics. Nevan Krogan has a financially compensated consulting agreement with Maze Therapeutics. Nevan is the President and is on the Board of Directors of Rezo Therapeutics, and he is a shareholder in Tenaya Therapeutics, Maze Therapeutics, Rezo Therapeutics, GEn1E Lifesciences, and Interline Therapeutics. J.S.F. is a consultant to, shareholder of, and receives sponsored research support from Relay Therapeutics. Michael Grabe is a developer for Berkeley Madonna.
Figures






References
-
- Backes Sandra, Hess Steffen, Boos Felix, Woellhaf Michael W., Gödel Sabrina, Jung Martin, Mühlhaus Timo, and Herrmann Johannes M.. 2018. “Tom70 Enhances Mitochondrial Preprotein Import Efficiency by Binding to Internal Targeting Sequences.” The Journal of Cell Biology 217 (4): 1369–82. - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous