

This is a preprint.
Altered auditory feature discrimination in a rat model of Fragile X Syndrome
- PMID: 40027738
- PMCID: PMC11870463
- DOI: 10.1101/2025.02.18.638956
Altered auditory feature discrimination in a rat model of Fragile X Syndrome
Update in
-
Altered auditory feature discrimination in a rat model of Fragile X Syndrome.PLoS Biol. 2025 Jul 1;23(7):e3003248. doi: 10.1371/journal.pbio.3003248. eCollection 2025 Jul. PLoS Biol. 2025. PMID: 40591708 Free PMC article.
Abstract
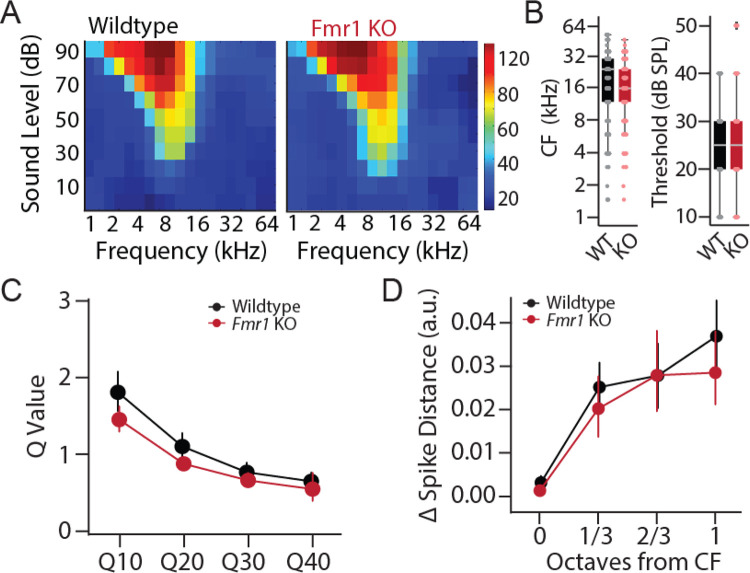
Atypical sensory processing, particularly in the auditory domain, is one of the most common and quality-of-life affecting symptoms seen in autism spectrum disorders (ASD). Fragile X Syndrome (FXS) is the leading inherited cause of ASD and a majority of FXS individuals present with auditory processing alterations. While auditory hypersensitivity is a common phenotype observed in FXS and Fmr1 KO rodent models, it is important to consider other auditory coding impairments that could contribute to sound processing difficulties and disrupted language comprehension in FXS. We have shown previously that a Fmr1 knockout (KO) rat model of FXS exhibits heightened sound sensitivity that coincided with abnormal perceptual integration of sound bandwidth, indicative of altered spectral processing. Frequency discrimination is a fundamental aspect of sound encoding that is important for a range of auditory processes, such as source segregation and speech comprehension, and disrupted frequency coding could thus contribute to a range of auditory issues in FXS and ASD. Here we explicitly characterized spectral processing deficits in male Fmr1 KO rats using an operant conditioning tone discrimination assay and in vivo electrophysiology recordings from the auditory cortex and inferior colliculus. We found that Fmr1 KO rats exhibited poorer frequency resolution, which corresponded with neuronal hyperactivity and broader frequency tuning in auditory cortical but not collicular neurons. Using an experimentally informed population model, we show that these cortical physiological differences can recapitulate the observed behavior discrimination deficits, with decoder performance being tightly linked to differences in cortical tuning width and signal-to-noise ratios. These findings suggest that cortical hyperexcitability may account for a range of auditory behavioral phenotypes in FXS, providing a potential locus for development of novel biomarkers and treatment strategies that could extend to other forms of ASD.
Keywords: auditory cortex; autism; fragile x syndrome; frequency discrimination; hyperexcitability; inferior colliculus; sensory processing; tuning.
Figures





References
-
- Hagerman R. J. et al. Fragile X syndrome. Nat. Rev. Dis. Primer 3, 1–19 (2017). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials