

Chemically Informed Coarse-Graining of Electrostatic Forces in Charge-Rich Biomolecular Condensates
- PMID: 40028356
- PMCID: PMC11869137
- DOI: 10.1021/acscentsci.4c01617
Chemically Informed Coarse-Graining of Electrostatic Forces in Charge-Rich Biomolecular Condensates
Abstract
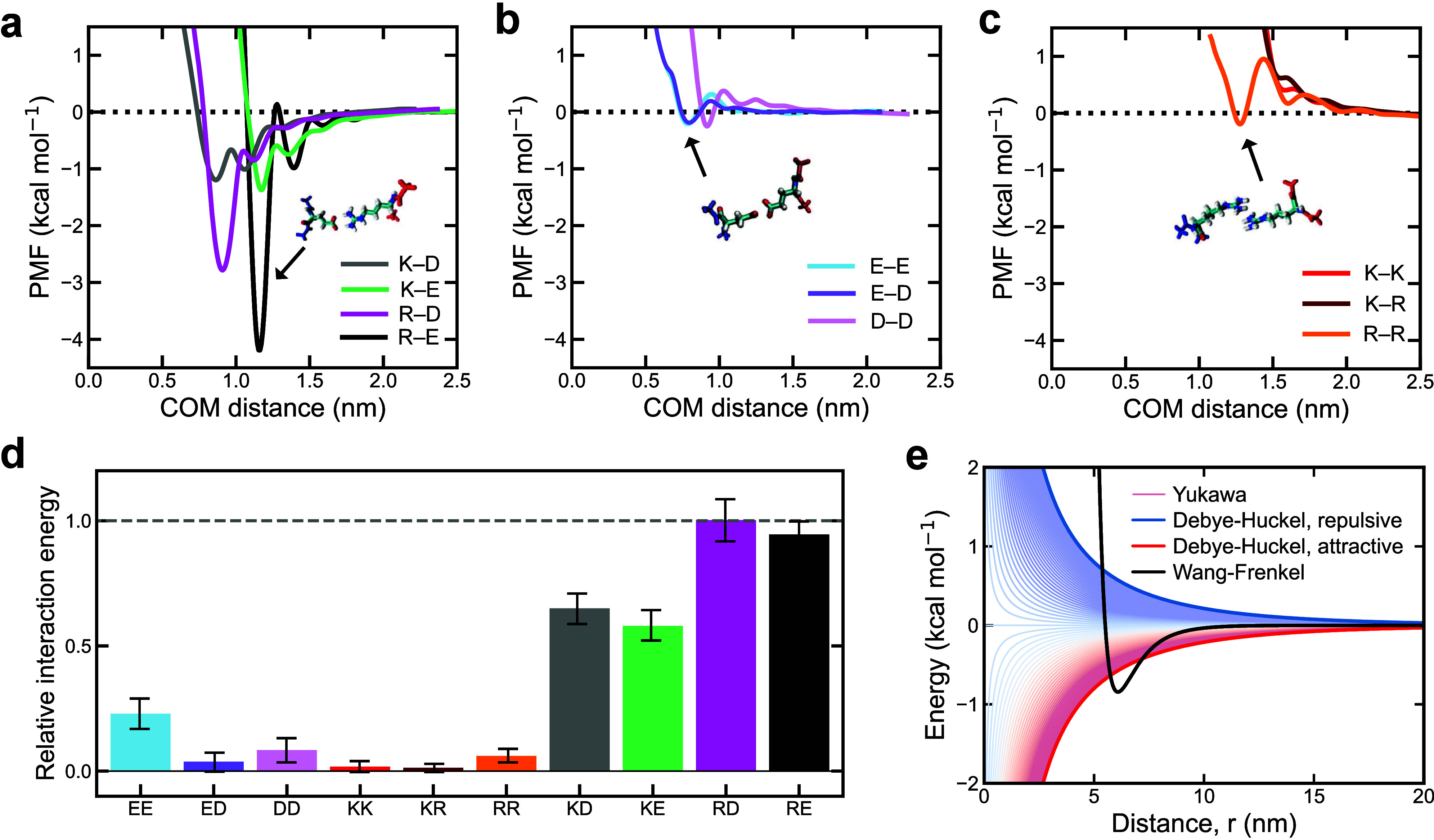
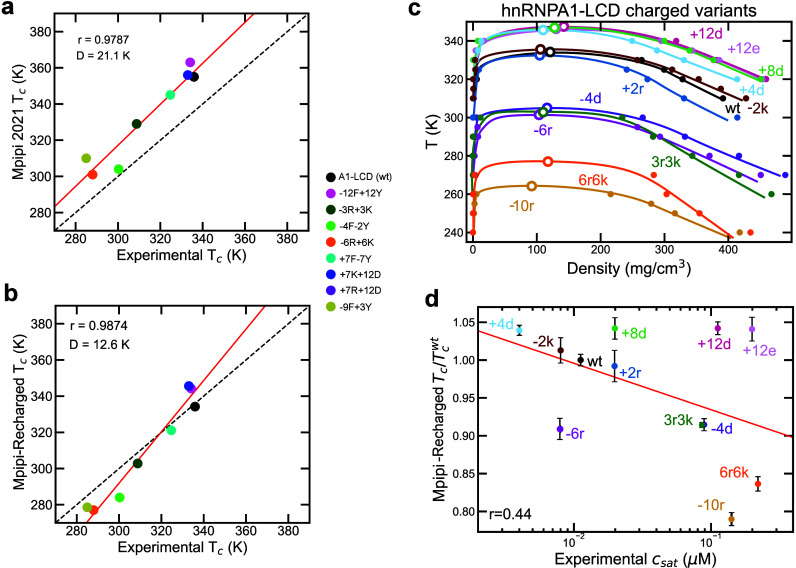
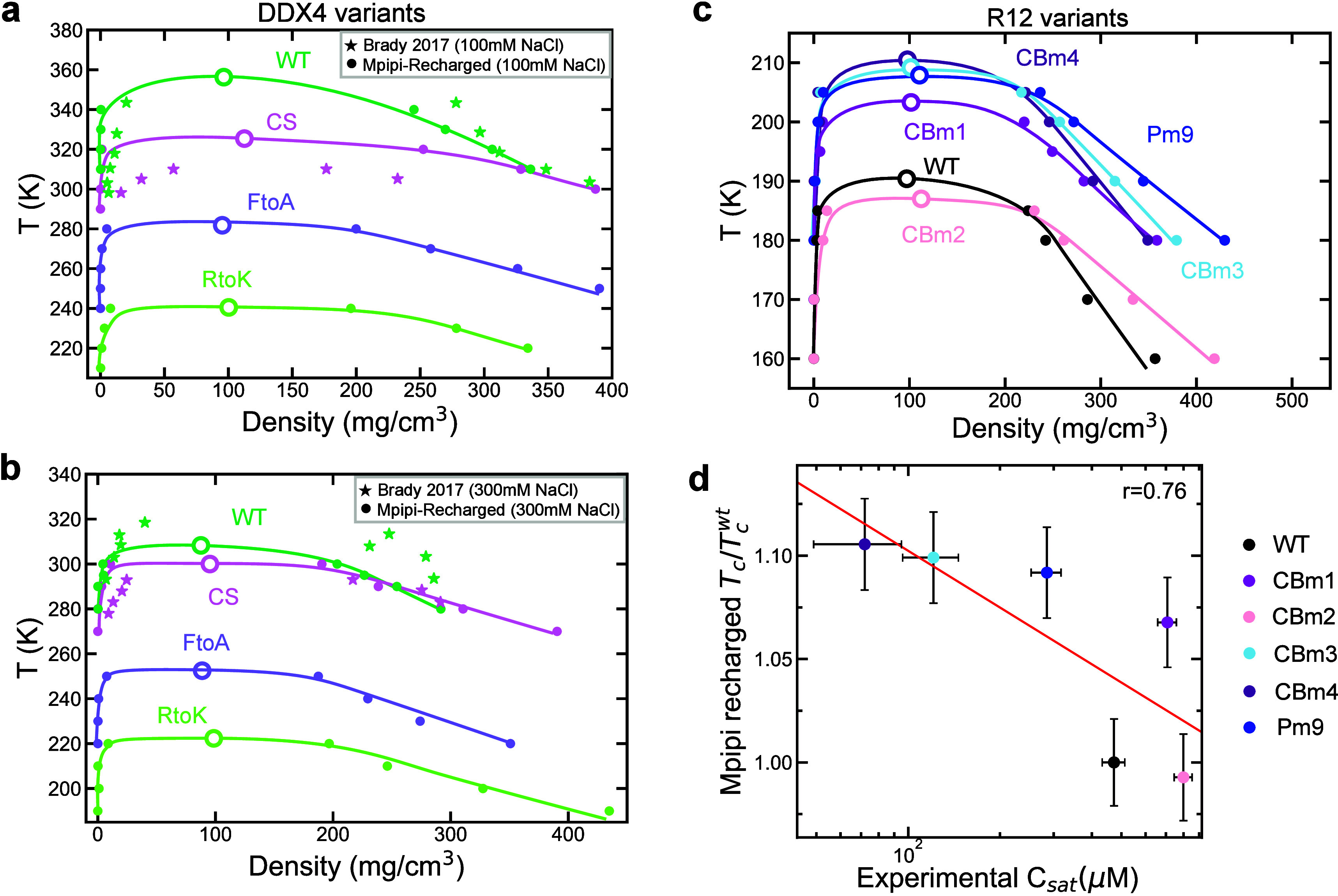
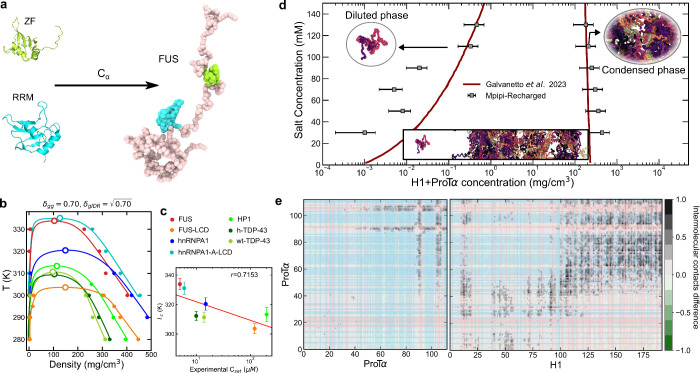
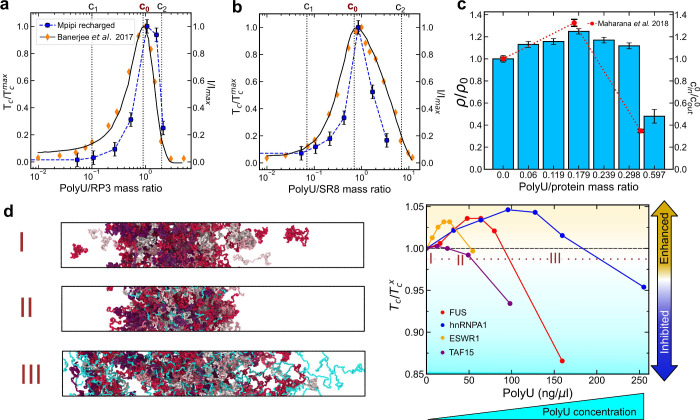
Biomolecular condensates composed of highly charged biomolecules, such as DNA, RNA, chromatin, and nucleic-acid binding proteins, are ubiquitous in the cell nucleus. The biophysical properties of these charge-rich condensates are largely regulated by electrostatic interactions. Residue-resolution coarse-grained models that describe solvent and ions implicitly are widely used to gain mechanistic insights into the biophysical properties of condensates, offering transferability, computational efficiency, and accurate predictions for multiple systems. However, their predictive accuracy diminishes for charge-rich condensates due to the implicit treatment of solvent and ions. Here, we present Mpipi-Recharged, a residue-resolution coarse-grained model that improves the description of charge effects in biomolecular condensates containing disordered proteins, multidomain proteins, and/or disordered single-stranded RNAs. Mpipi-Recharged introduces a pair-specific asymmetric Yukawa electrostatic potential, informed by atomistic simulations. We show that this asymmetric coarse-graining of electrostatic forces captures intricate effects, such as charge blockiness, stoichiometry variations in complex coacervates, and modulation of salt concentration, without requiring explicit solvation. Mpipi-Recharged provides excellent agreement with experiments in predicting the phase behavior of highly charged condensates. Overall, Mpipi-Recharged improves the computational tools available to investigate the physicochemical mechanisms regulating biomolecular condensates, enhancing the scope of computer simulations in this field.
© 2025 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Similar articles
-
Benchmarking residue-resolution protein coarse-grained models for simulations of biomolecular condensates.PLoS Comput Biol. 2025 Jan 13;21(1):e1012737. doi: 10.1371/journal.pcbi.1012737. eCollection 2025 Jan. PLoS Comput Biol. 2025. PMID: 39804953 Free PMC article.
-
Decoding phase separation of prion-like domains through data-driven scaling laws.Elife. 2025 Feb 12;13:RP99068. doi: 10.7554/eLife.99068. Elife. 2025. PMID: 39937084 Free PMC article.
-
Coarse-Grained Model of Disordered RNA for Simulations of Biomolecular Condensates.J Chem Theory Comput. 2025 Mar 11;21(5):2766-2779. doi: 10.1021/acs.jctc.4c01646. Epub 2025 Feb 26. J Chem Theory Comput. 2025. PMID: 40009520
-
Toward Predictive Coarse-Grained Simulations of Biomolecular Condensates.Biochemistry. 2025 Apr 15;64(8):1750-1761. doi: 10.1021/acs.biochem.4c00737. Epub 2025 Apr 2. Biochemistry. 2025. PMID: 40172489 Review.
-
Deciphering driving forces of biomolecular phase separation from simulations.Curr Opin Struct Biol. 2025 Jun;92:103026. doi: 10.1016/j.sbi.2025.103026. Epub 2025 Mar 8. Curr Opin Struct Biol. 2025. PMID: 40058249 Review.
Cited by
-
Controlled liquid-liquid phase separation via the simulation-guided, targeted engineering of the RNA-binding protein PARCL.iScience. 2025 Jun 11;28(7):112852. doi: 10.1016/j.isci.2025.112852. eCollection 2025 Jul 18. iScience. 2025. PMID: 40662195 Free PMC article.
References
-
- Espinosa J. R.; Joseph J. A.; Sanchez-Burgos I.; Garaizar A.; Frenkel D.; Collepardo-Guevara R. Liquid network connectivity regulates the stability and composition of biomolecular condensates with many components. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 13238–13247. 10.1073/pnas.1917569117. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources