

Broken beyond repair: TA system ParE toxins mediate effective gyrase inhibition without driving resistance
- PMID: 40029095
- PMCID: PMC11925232
- DOI: 10.1128/jb.00416-24
Broken beyond repair: TA system ParE toxins mediate effective gyrase inhibition without driving resistance
Abstract
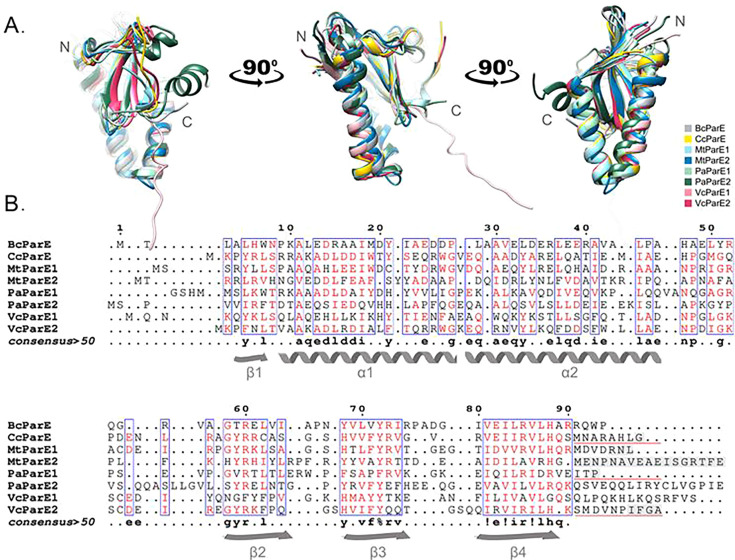
DNA gyrase is an essential bacterial-specific type IIA topoisomerase that corrects DNA overwinding during transcription and replication. Compounds capable of stabilizing gyrase-mediated double-strand DNA breaks are valuable antibacterials; however, these can trigger error-prone repair, potentially inducing DNA mutations leading to antimicrobial resistance. ParE toxin proteins, which belong to a family of type II toxin-antitoxin systems, inhibit DNA gyrase and promote the persistence of double-strand DNA breaks. However, it is unclear if the ParE-induced gyrase inhibition is equivalent for all ParE family members, or if any mutations arise and can accumulate to cause antibiotic resistance. Selected chromosomal ParE toxins were examined for toxicity to their native bacterial hosts, and the frequency of mutations and impact on susceptibility to selected antibiotics were assessed. Our results show that ParE toxins from Burkholderia cenocepacia, Mycobacterium tuberculosis, Pseudomonas aeruginosa, and Vibrio cholerae exert potent toxicities toward the native cells, whereas one tested ParE toxin from P. aeruginosa was not toxic. The contribution to toxicity of the ParE toxin C-terminal amino acid sequences was examined using two lab-generated chimeric ParE toxins; our results demonstrate that this region did not impact the toxicity level. Our study finds that the relative potency of individual ParE toxins correlates with increases in mutation frequency. While some ParE toxins induced limited collateral sensitivity to selected antibiotics, no increases in MIC values were found. Overall, this study demonstrates the relative toxicity of different ParE toxins. Importantly, the toxicity appears to result in loss of viability before productive resistance-inducing mutations can accumulate.
Importance: Toxin-antitoxin (TA) systems can halt growth or kill cells when the toxin protein engages with the host cell target. In the ParDE TA system, the toxin ParE inhibits DNA gyrase, resulting in loss of viability that phenocopies fluoroquinolone antibiotics. Our study demonstrates that ParE toxins increase the frequency of mutations, presumably by a mechanism similar to fluoroquinolone antibiotics. These increases scale to the resulting toxicity, and importantly, these mutations do not accumulate into productive antibacterial resistance. This suggests that ParE toxins are not intrinsic drivers of resistance and, if the molecular mechanism can be harnessed, could generate a new class of gyrase inhibitors.
Keywords: ParE toxin; antimicrobial susceptibility; bacterial cell viability; gyrase inhibition; mutation frequency; toxin antitoxin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
