

This is a preprint.
Three-dimensional regulatory hubs support oncogenic programs in glioblastoma
- PMID: 40034649
- PMCID: PMC11875237
- DOI: 10.1101/2024.12.20.629544
Three-dimensional regulatory hubs support oncogenic programs in glioblastoma
Update in
-
Three-dimensional regulatory hubs support oncogenic programs in glioblastoma.Mol Cell. 2025 Apr 3;85(7):1330-1348.e6. doi: 10.1016/j.molcel.2025.03.007. Epub 2025 Mar 26. Mol Cell. 2025. PMID: 40147440
Abstract
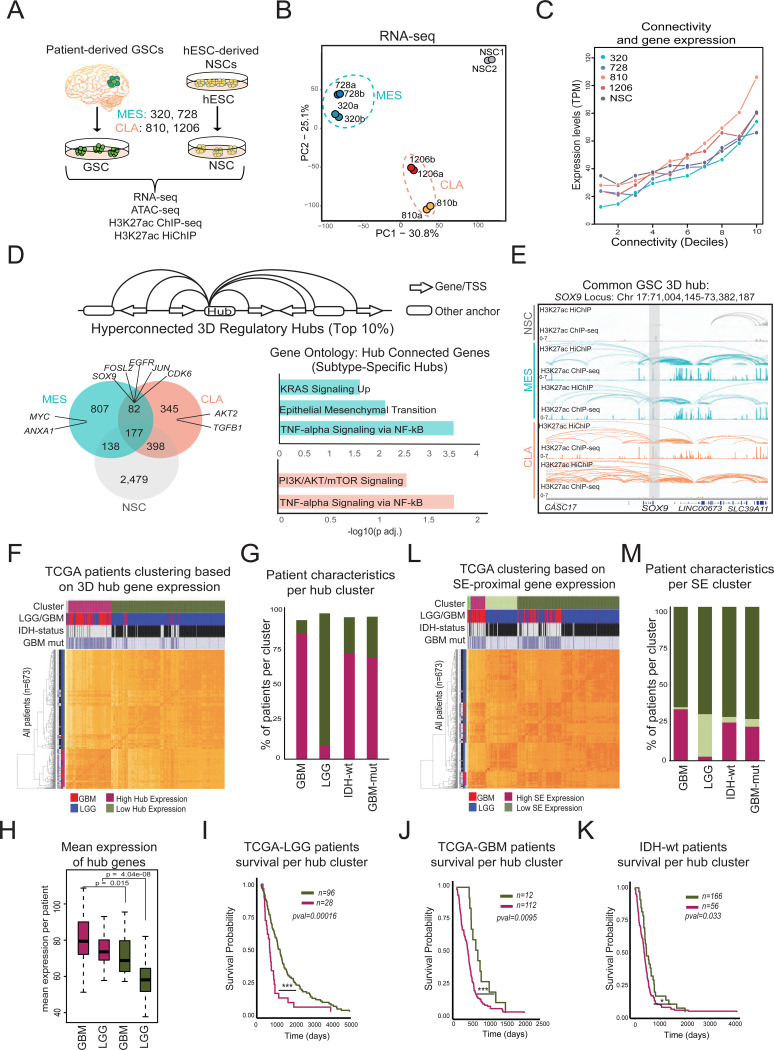
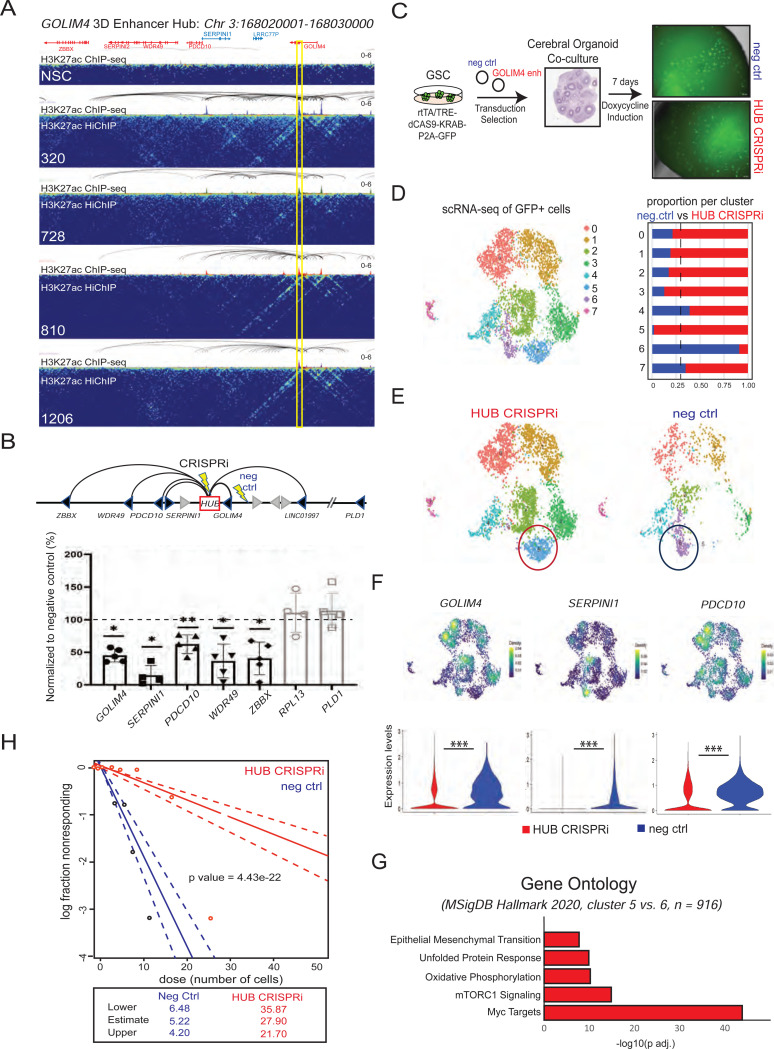
Dysregulation of enhancer-promoter communication in the context of the three-dimensional (3D) nucleus is increasingly recognized as a potential driver of oncogenic programs. Here, we profiled the 3D enhancer-promoter networks of primary patient-derived glioblastoma stem cells (GSCs) in comparison with neuronal stem cells (NSCs) to identify potential central nodes and vulnerabilities in the regulatory logic of this devastating cancer. Specifically, we focused on hyperconnected 3D regulatory hubs and demonstrated that hub-interacting genes exhibit high and coordinated expression at the single-cell level and strong association with oncogenic programs that distinguish IDH-wt glioblastoma patients from low-grade glioma. Epigenetic silencing of a recurrent 3D enhancer hub-with an uncharacterized role in glioblastoma-was sufficient to cause concordant downregulation of multiple hub-connected genes along with significant shifts in transcriptional states and reduced clonogenicity. By integrating published datasets from other cancer types, we also identified both universal and cancer type-specific 3D regulatory hubs which enrich for varying oncogenic programs and nominate specific factors associated with worse outcomes. Genetic alterations, such as focal duplications, could explain only a small fraction of the detected hyperconnected hubs and their increased activity. Overall, our study provides computational and experimental support for the potential central role of 3D regulatory hubs in controlling oncogenic programs and properties.
Keywords: 3D chromatin organization; CRISPRi; HiChIP; clonogenicity; enhancer hubs; enhancer-promoter interactions; glioblastoma; oncogenic program; single-cell RNA-seq; structural variants.
Conflict of interest statement
Conflict of interest statement The authors declare that the above study was conducted in the absence of any commercial, financial, or personal relationships that could have appeared to influence the work reported in this article. All authors have approved the submitted version.
Figures


References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources