

Mug20-Rec25-Rec27 binds DNA and enhances meiotic DNA break formation via phase-separated condensates
- PMID: 40037704
- PMCID: PMC11879393
- DOI: 10.1093/nar/gkaf123
Mug20-Rec25-Rec27 binds DNA and enhances meiotic DNA break formation via phase-separated condensates
Abstract
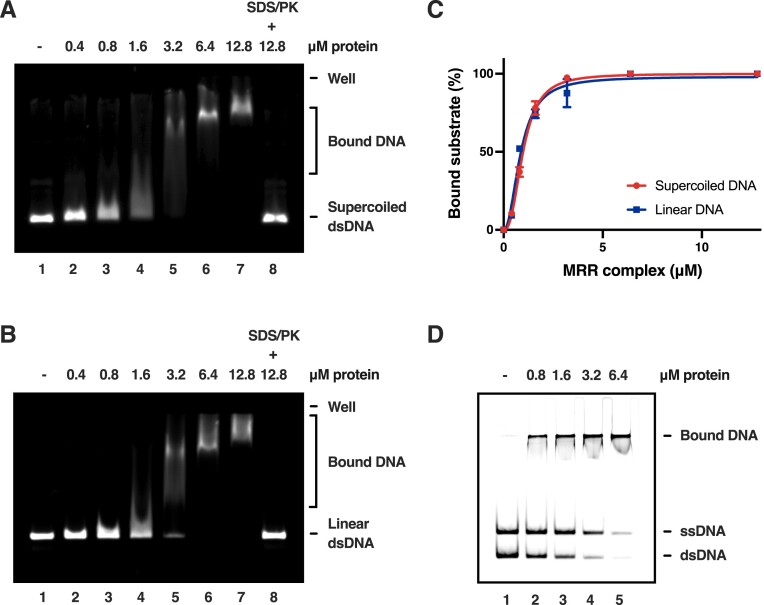
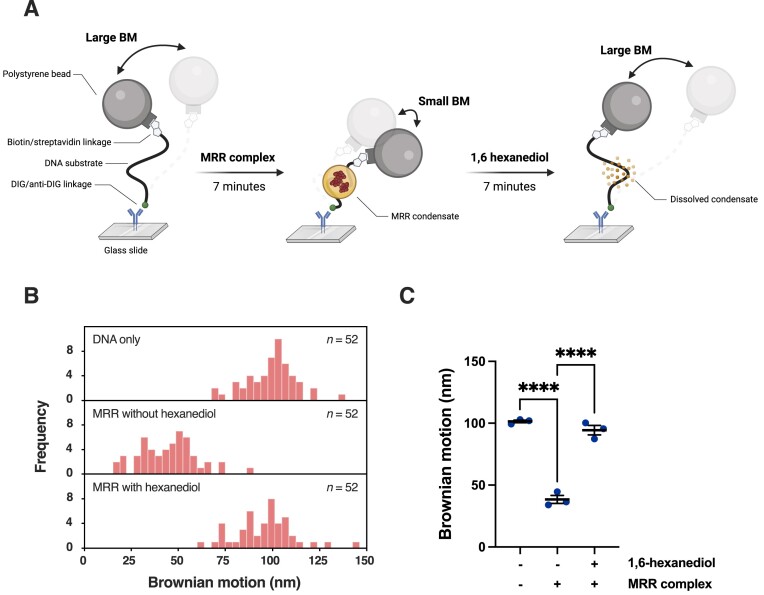
During meiosis, programmed DNA double-strand breaks (DSBs) are formed at hotspots to initiate homologous recombination, which is vital for reassorting genetic material. In fission yeast, the linear element (LinE) proteins Mug20, Rec25, and Rec27 interdependently bind chromosomal hotspots with high specificity and are necessary for high-level DSB formation. However, their mechanistic role in regulating the meiotic DSB machinery remains unknown. Here, using purified Mug20-Rec25-Rec27 (MRR) complex and functional intracellular analyses, we reveal that the MRR-DNA nucleoprotein complex assembles phase-separated condensates that compact the DNA. Notably, MRR complex formation is a prerequisite for DNA binding and condensate assembly, with Rec27 playing a pivotal role in directly binding DNA. Consistent with this finding, failure to form MRR-DNA condensates results in defective intracellular meiotic DSB formation and recombination. Our results provide mechanistic insights into how LinEs enhance meiotic DSB formation and provide a paradigm for studies in other species.
© The Author(s) 2025. Published by Oxford University Press on behalf of Nucleic Acids Research.
Conflict of interest statement
The authors declare no competing interests.
Figures






References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
