

Regeneration in planarians modifies behavioral switching
- PMID: 40040806
- PMCID: PMC11879593
- DOI: 10.1016/j.isci.2025.111947
Regeneration in planarians modifies behavioral switching
Abstract
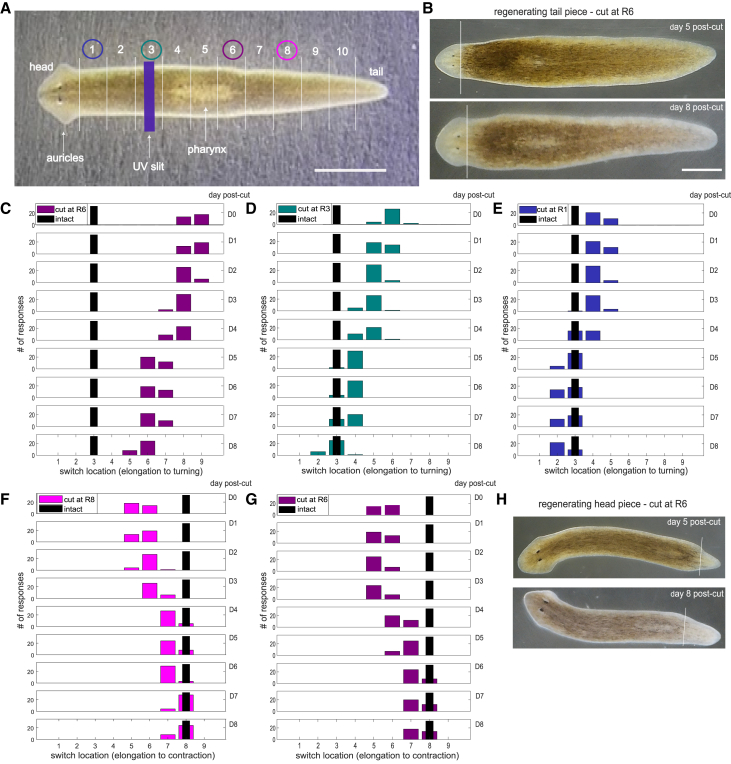
The planarian Dugesia japonica responds differently to localized stimuli: anterior regions turn, middle regions elongate, and posterior regions contract. If cut into several pieces, each piece immediately produces the same three responses. Over several days, each piece regenerates all transected body parts. This study tested how the pieces coordinate behavioral responses during regeneration. We first determined the locations of the turning/elongation and elongation/contraction behavioral switches. Immediately, all transections moved both switching sites away from the cut sites so that the worm pieces produced the same three responses as intact worms. During regeneration, the sites of behavioral switching moved progressively closer to the transection (now regeneration) sites. These results show that the immediate effects of transection (likely physiological) are coordinated with the addition of regenerating tissue (anatomical) to maintain as normal an animal as possible. Other animals that regenerate body parts, such as amphibians and reptiles, may use similar coordination mechanisms.
Keywords: Biological sciences; Developmental anatomy; Histology.
© 2025 The Authors. Published by Elsevier Inc.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Grants and funding
LinkOut - more resources
Full Text Sources
