

Immunomodulatory role of the stem cell circadian clock in muscle repair
- PMID: 40043110
- PMCID: PMC11881903
- DOI: 10.1126/sciadv.adq8538
Immunomodulatory role of the stem cell circadian clock in muscle repair
Abstract
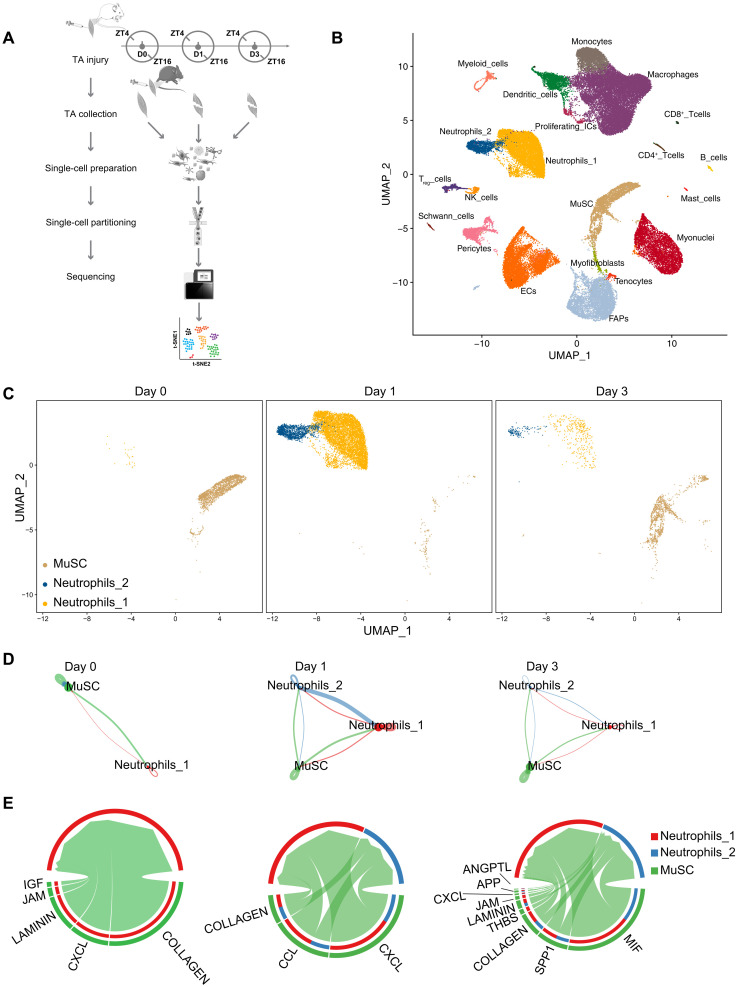
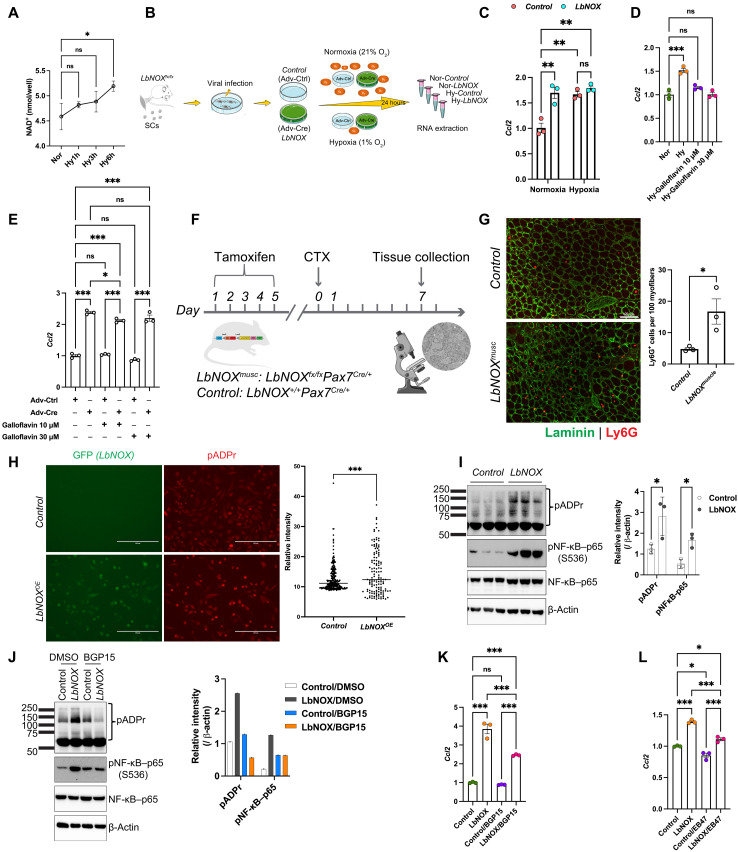
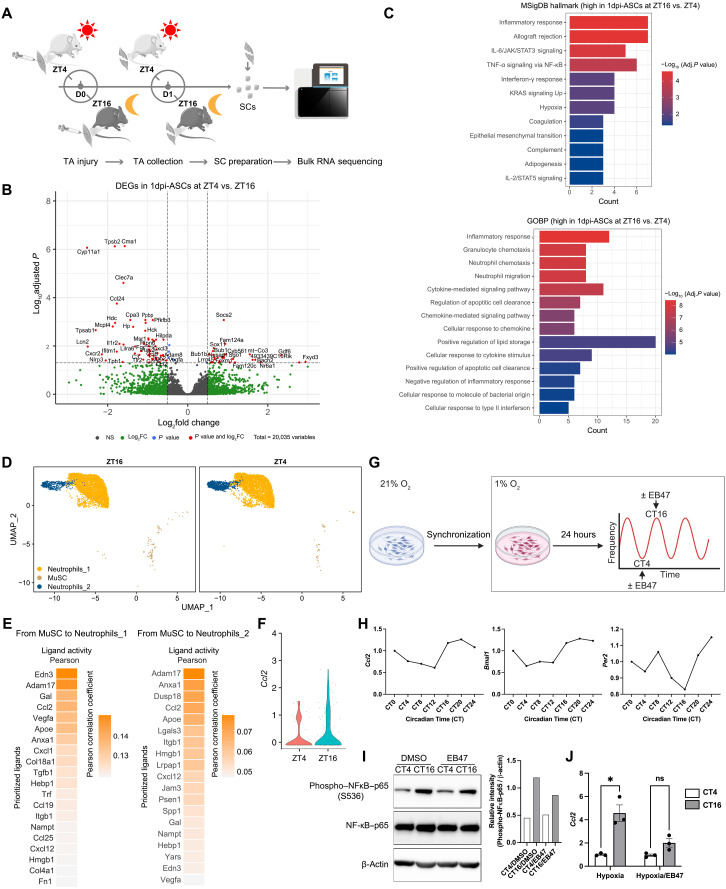
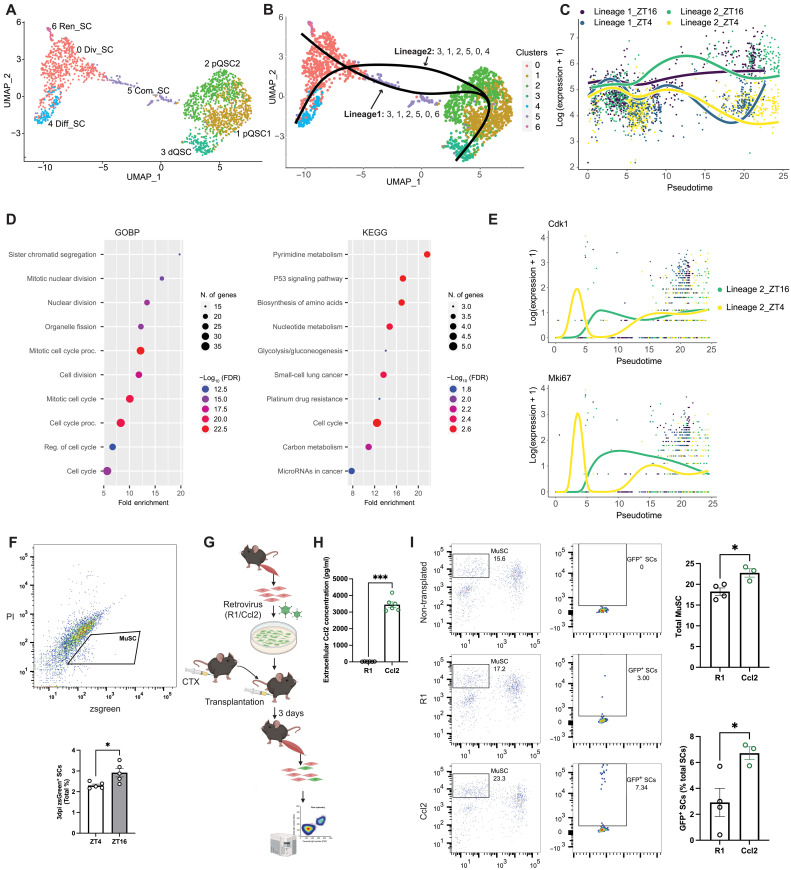
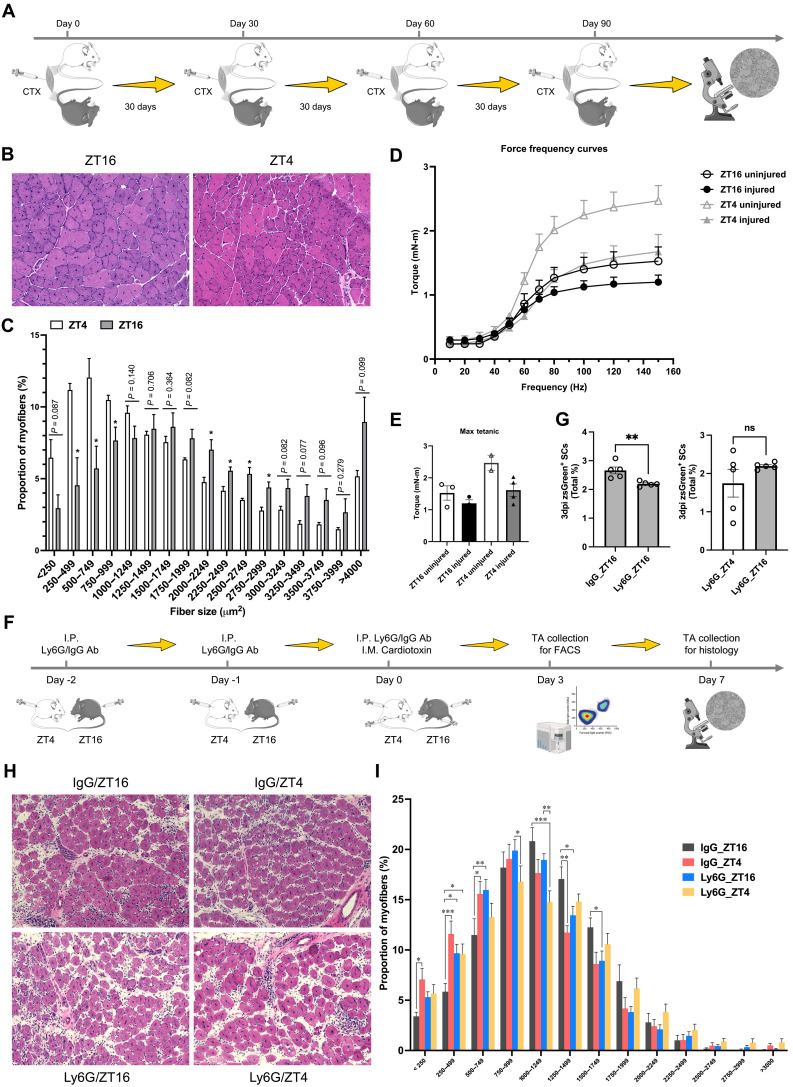
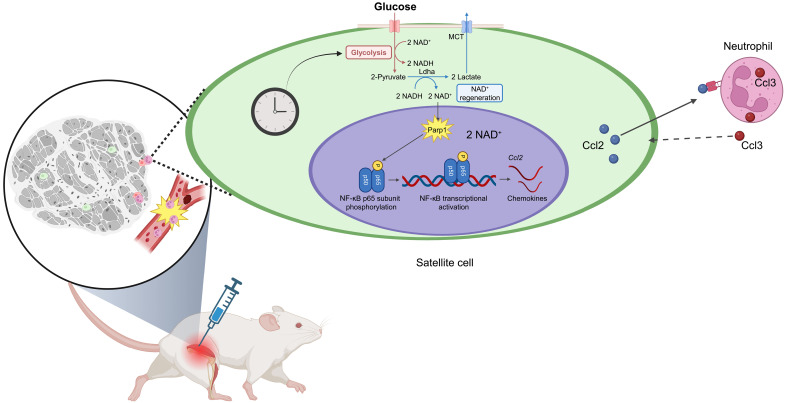
Circadian rhythms orchestrate physiological processes such as metabolism, immune function, and tissue regeneration, aligning them with the optimal time of day (TOD). This study identifies an interplay between the circadian clock within muscle stem cells (SCs) and their capacity to modulate the immune microenvironment during muscle regeneration. We reveal that the SC clock triggers TOD-dependent inflammatory gene transcription after injury, particularly genes related to neutrophil activity and chemotaxis. These responses are driven by cytosolic regeneration of the signaling metabolite nicotinamide adenine dinucleotide (oxidized form) (NAD+), as enhancing cytosolic NAD+ regeneration in SCs is sufficient to induce inflammatory responses that influence muscle regeneration. Mononuclear single-cell sequencing of the regenerating muscle niche further implicates the cytokine CCL2 in mediating SC-neutrophil cross-talk in a TOD-dependent manner. Our findings highlight the intersection between SC metabolic shifts and immune responses within the muscle microenvironment, dictated by circadian rhythms, and underscore the potential for targeting circadian and metabolic pathways to enhance tissue regeneration.
Figures

Update of
-
Immunomodulatory Role of the Stem Cell Circadian Clock in Muscle Repair.bioRxiv [Preprint]. 2024 May 28:2024.05.24.595728. doi: 10.1101/2024.05.24.595728. bioRxiv. 2024. Update in: Sci Adv. 2025 Mar 07;11(10):eadq8538. doi: 10.1126/sciadv.adq8538. PMID: 38854114 Free PMC article. Updated. Preprint.
References
-
- Bass J., Lazar M. A., Circadian time signatures of fitness and disease. Science 354, 994–999 (2016). - PubMed
-
- Zhu P., Hamlish N. X., Thakkar A. V., Steffeck A. W. T., Rendleman E. J., Khan N. H., Waldeck N. J., DeVilbiss A. W., Martin-Sandoval M. S., Mathews T. P., Chandel N. S., Peek C. B., BMAL1 drives muscle repair through control of hypoxic NAD+ regeneration in satellite cells. Genes Dev. 36, 149–166 (2022). - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
