

Solanum pan-genetics reveals paralogues as contingencies in crop engineering
- PMID: 40044854
- PMCID: PMC11964936
- DOI: 10.1038/s41586-025-08619-6
Solanum pan-genetics reveals paralogues as contingencies in crop engineering
Abstract
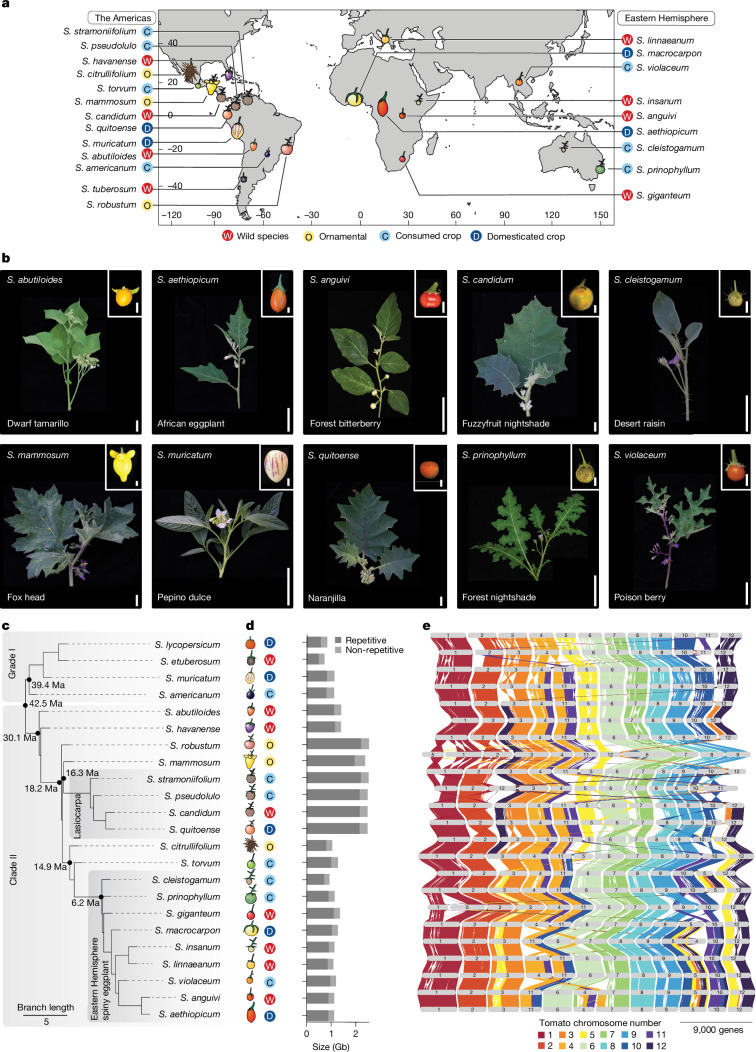
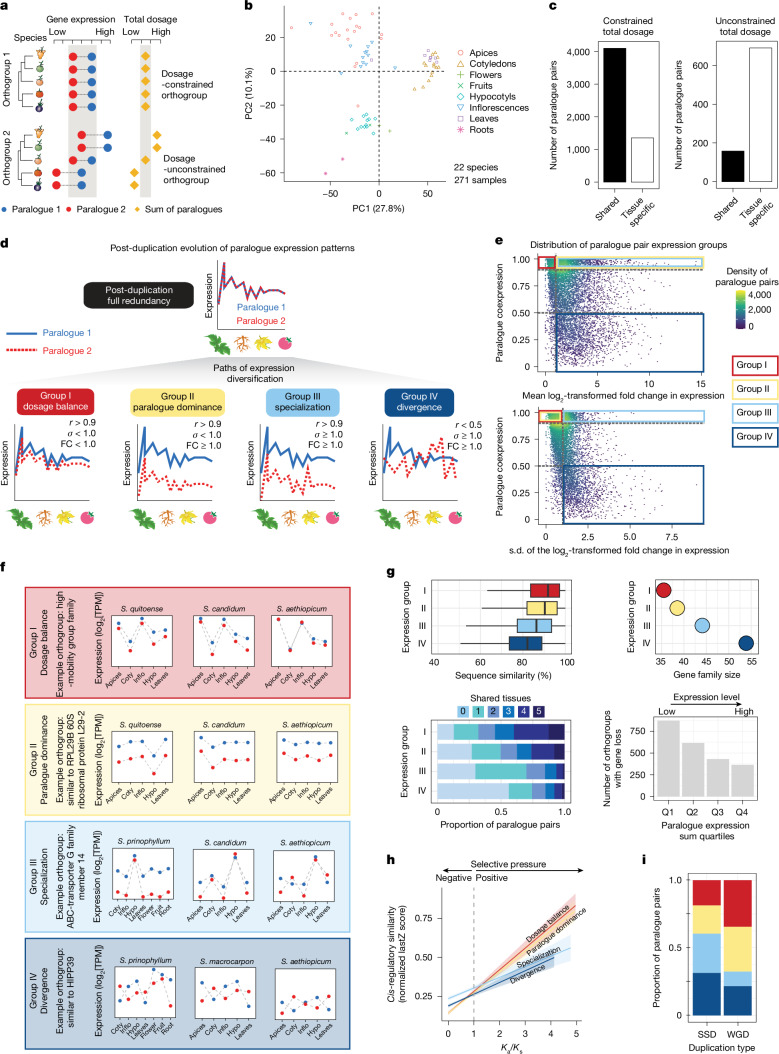
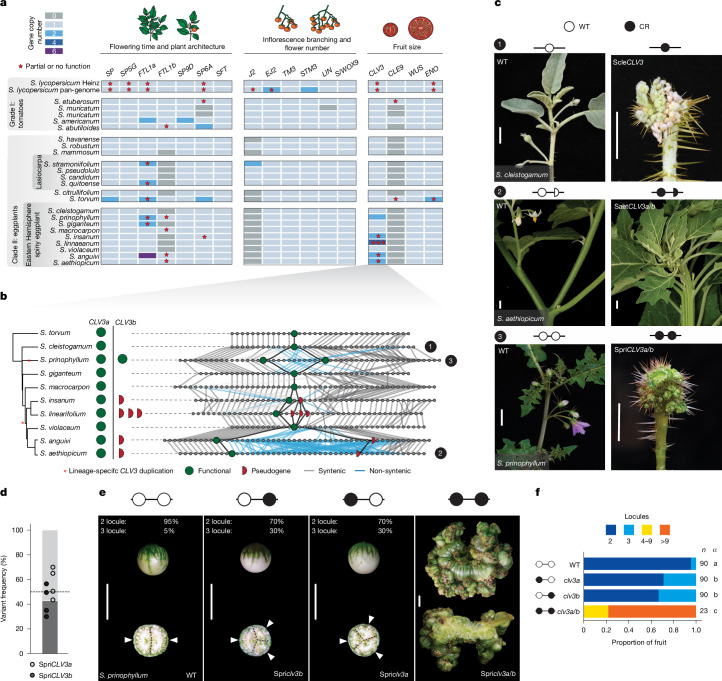
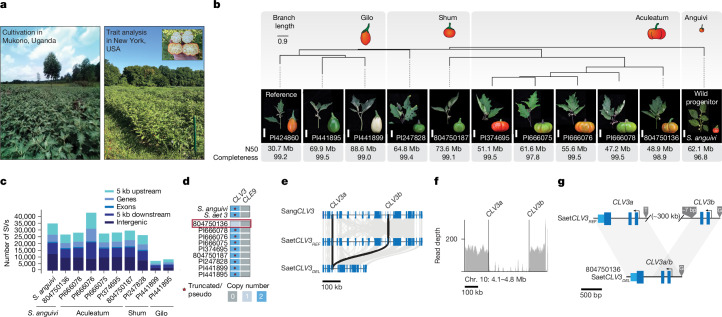
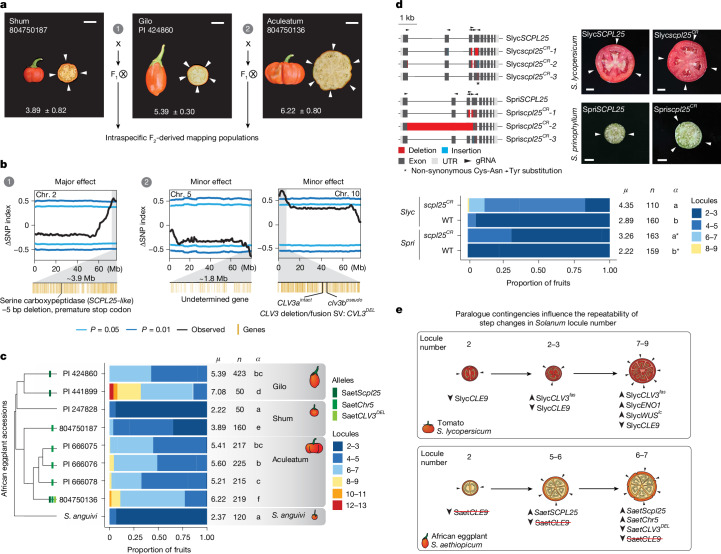
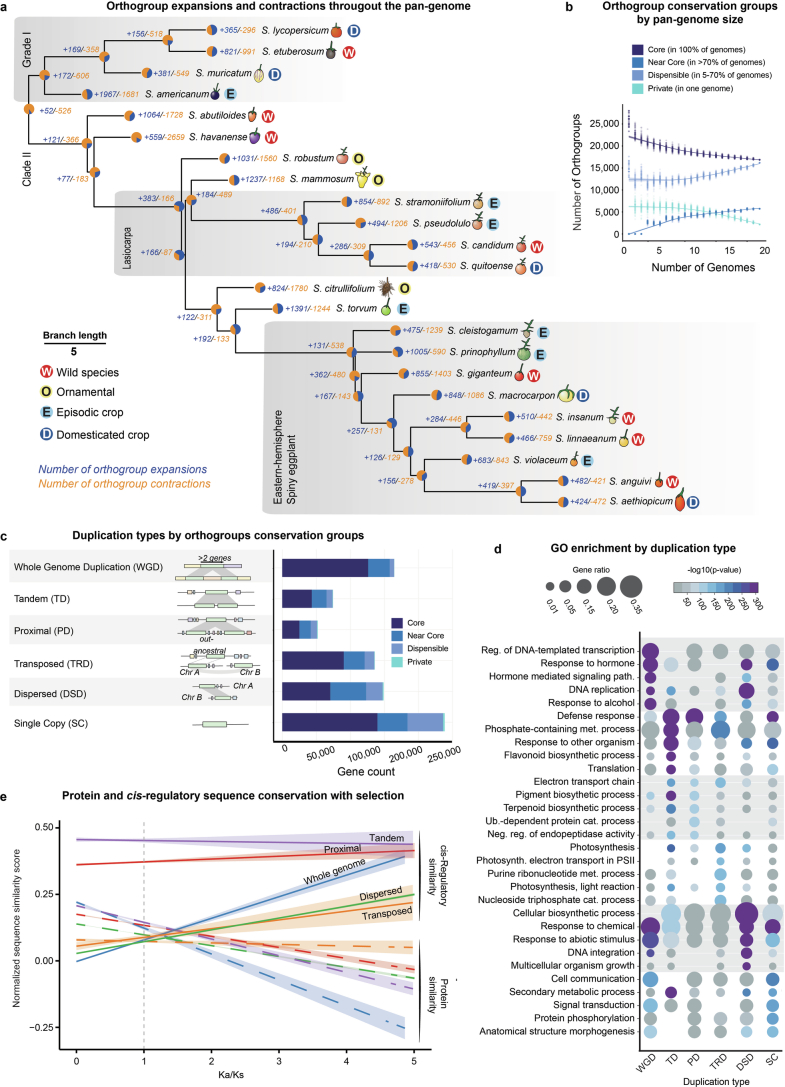
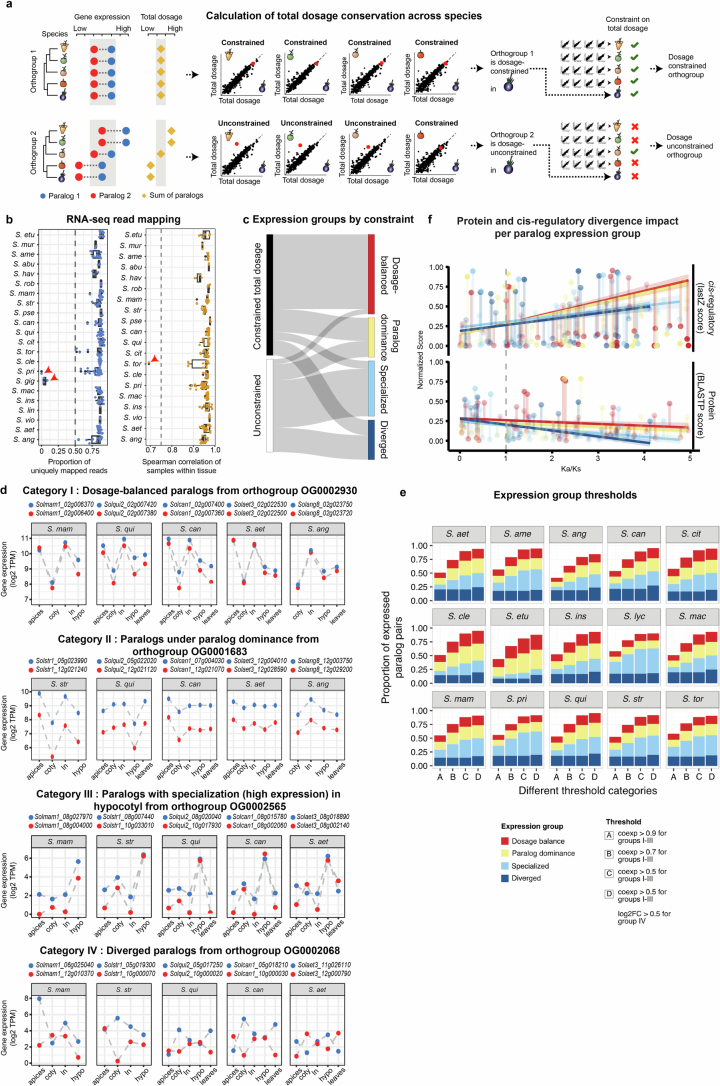
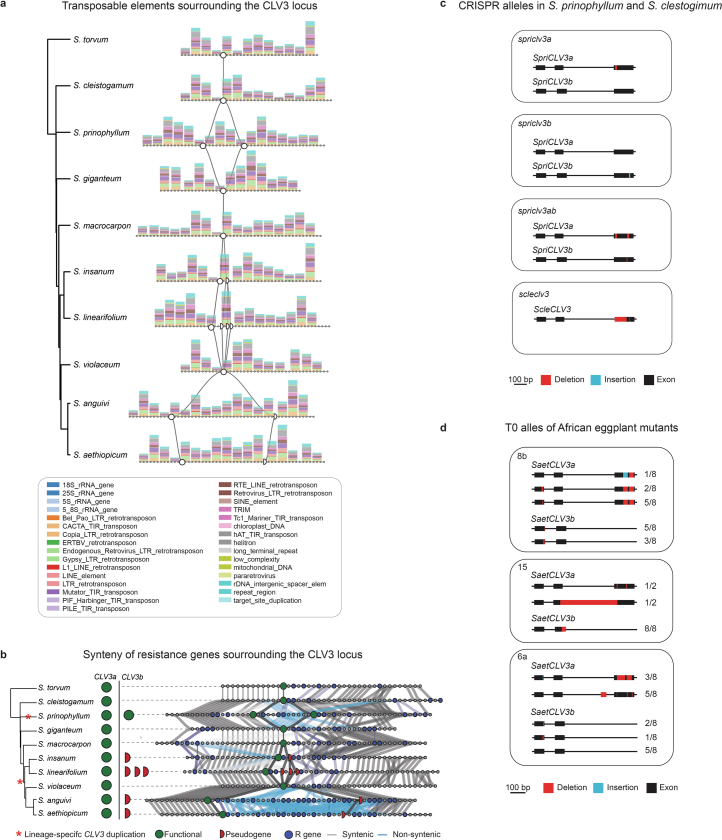
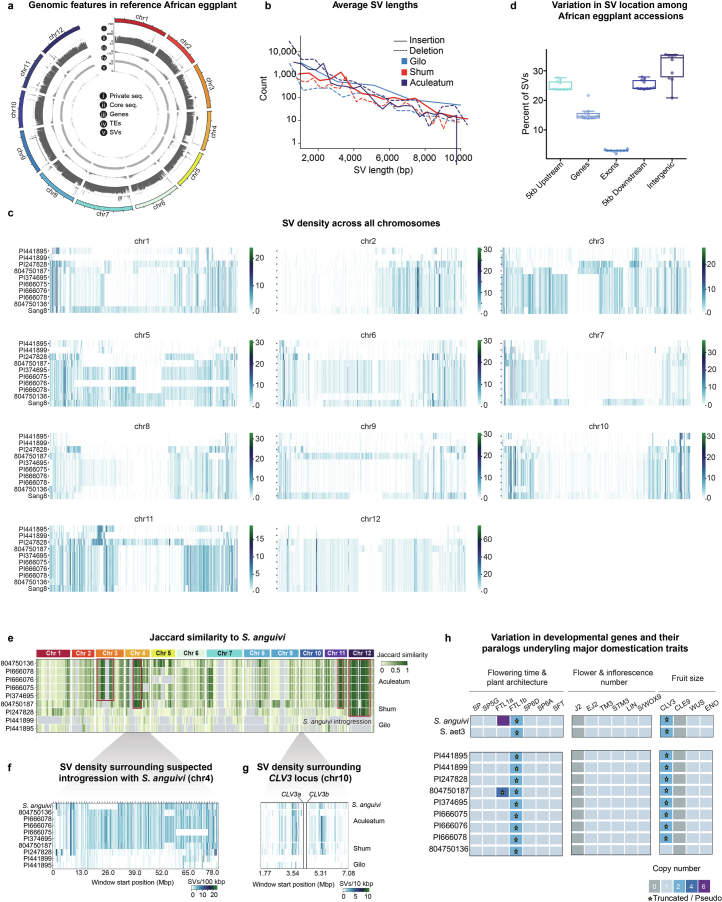
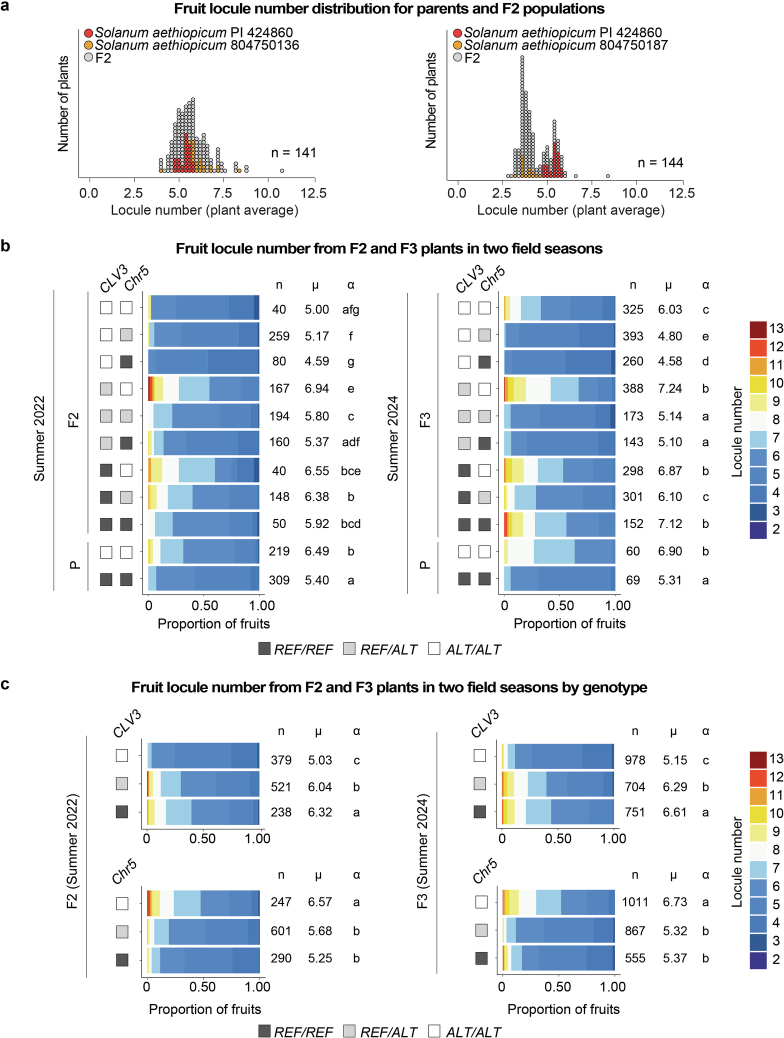
Pan-genomics and genome-editing technologies are revolutionizing breeding of global crops1,2. A transformative opportunity lies in exchanging genotype-to-phenotype knowledge between major crops (that is, those cultivated globally) and indigenous crops (that is, those locally cultivated within a circumscribed area)3-5 to enhance our food system. However, species-specific genetic variants and their interactions with desirable natural or engineered mutations pose barriers to achieving predictable phenotypic effects, even between related crops6,7. Here, by establishing a pan-genome of the crop-rich genus Solanum8 and integrating functional genomics and pan-genetics, we show that gene duplication and subsequent paralogue diversification are major obstacles to genotype-to-phenotype predictability. Despite broad conservation of gene macrosynteny among chromosome-scale references for 22 species, including 13 indigenous crops, thousands of gene duplications, particularly within key domestication gene families, exhibited dynamic trajectories in sequence, expression and function. By augmenting our pan-genome with African eggplant cultivars9 and applying quantitative genetics and genome editing, we dissected an intricate history of paralogue evolution affecting fruit size. The loss of a redundant paralogue of the classical fruit size regulator CLAVATA3 (CLV3)10,11 was compensated by a lineage-specific tandem duplication. Subsequent pseudogenization of the derived copy, followed by a large cultivar-specific deletion, created a single fused CLV3 allele that modulates fruit organ number alongside an enzymatic gene controlling the same trait. Our findings demonstrate that paralogue diversifications over short timescales are underexplored contingencies in trait evolvability. Exposing and navigating these contingencies is crucial for translating genotype-to-phenotype relationships across species.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: W.R.M. is a founder and shareholder in Orion Genomics, a plant genomics company. Z.B.L. is a consultant for and a member of the Scientific Strategy Board of Inari Agriculture. The other authors declare no competing interests.
Figures
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
