

Genetic and pharmacologic enhancement of SUMO2 conjugation prevents and reverses cognitive impairment and synaptotoxicity in a preclinical model of Alzheimer's disease
- PMID: 40047257
- PMCID: PMC11883658
- DOI: 10.1002/alz.70030
Genetic and pharmacologic enhancement of SUMO2 conjugation prevents and reverses cognitive impairment and synaptotoxicity in a preclinical model of Alzheimer's disease
Erratum in
-
Correction to "Genetic and pharmacologic enhancement of SUMO2 conjugation prevents and reverses cognitive impairment and synaptotoxicity in a preclinical model of Alzheimer's disease".Alzheimers Dement. 2025 Aug;21(8):e70606. doi: 10.1002/alz.70606. Alzheimers Dement. 2025. PMID: 40847048 Free PMC article. No abstract available.
Abstract
Introduction: Amyloid beta oligomers (Aβos) are toxic to synapses and key to the progression of Alzheimer's disease (AD) and amyloid pathology, representing a target for therapeutic strategies.
Methods: Amyloid and small ubiquitin modifier 2 (SUMO2) transgenics were analyzed by electrophysiology and behavioral testing. A recombinant analogue of SUMO2, SBT02, was generated and assessed for brain penetration and the ability to mitigate amyloid pathology.
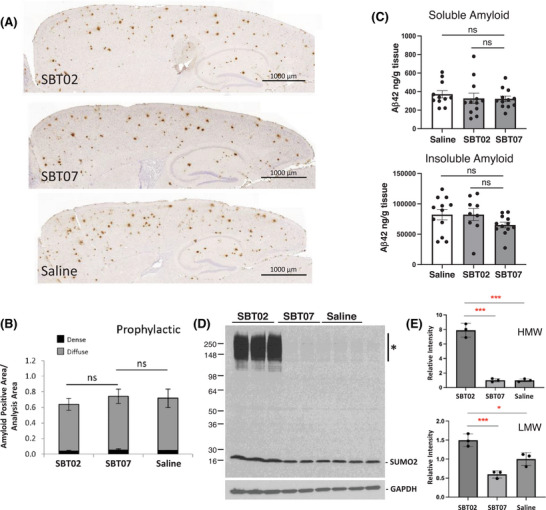
Results: Elevated SUMO2 expression prevents cognitive and synaptic impairment in a mouse model of AD amyloid pathology. Systemic administration of SBT02 resulted in high brain bioavailability and prophylactically halted the progression of AD-associated deficits. SBT02 also restored cognition and synaptic function in late-stage amyloid load. Mechanistically, SUMO2 and SBT02 do not alter amyloid processing or clearance and mitigate synaptotoxicity in the presence of high amyloid loads.
Discussion: SBT02 is a promising therapeutic strategy to counteract and reverse the toxic effects of Aβos in AD.
Highlights: Genetic overexpression of human SUMO2 prevents the long-term potentiation (LTP) impairments and cognitive deficits in amyloid precursor protein (APP) transgenics without affecting amyloid pathology. A recombinant analogue of human SUMO2, termed SBT02, when administered systemically, displays high brain bioavailability and has no adverse effects at high doses. Prophylactic treatment of APP transgenics with SBT02 prior to the development of amyloid pathology results in the prevention of synaptic and behavioral dysfunction. SBT02 also reverses pre-existing LTP and cognitive impairments when administered to APP transgenics with advanced and severe pathology. SBT02 has no impact on amyloid pathology, indicating a mechanism of action on synaptic resistance to Aβ toxicity.
Keywords: SUMOylation; amyloid transgenic mouse model; behavioral analyses; biologic therapy; cognition; drug development; electrophysiology; preclinical testing; small ubiquitin‐like modifiers.
© 2025 The Author(s). Alzheimer's & Dementia published by Wiley Periodicals LLC on behalf of Alzheimer's Association.
Conflict of interest statement
L.F., O.A., and P.E.F. are co‐inventors on a patent application for the use of SUMO2 mimetic peptides in Alzheimer's disease. N.W., E.K.A., S.M., H.T., K.S., K.H., H.Y., A.S., E.A., F.O., A.M., and T.K. have nothing to disclose. Author disclosures are available in the Supporting Information.
Figures







References
MeSH terms
Substances
Grants and funding
- NIH-NINDS R01NS110024/NH/NIH HHS/United States
- R01 NS110024/NS/NINDS NIH HHS/United States
- 17KK0197 to S.M/Fostering Joint International Research
- NIH R01NS134902 to L.F/NH/NIH HHS/United States
- R01NS049442 to O.A/NH/NIH HHS/United States
- R01 NS134902/NS/NINDS NIH HHS/United States
- AARG 17-505136 to L.F/ALZ/Alzheimer's Association/United States
- Japanese Society for the Promotion of Science
- PJT-173497/CAPMC/ CIHR/Canada
- TR192065/Weston Brain Institute
- 21K06759 to S.M/Grant-in-Aid for Scientific Research
- Japanese Society for the Promotion of Science (JSPS)
- R01 NS049442/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
