

Another tail of two sites: activation of the Notch ligand Delta by Mindbomb1
- PMID: 40050848
- PMCID: PMC11887331
- DOI: 10.1186/s12915-025-02162-6
Another tail of two sites: activation of the Notch ligand Delta by Mindbomb1
Abstract
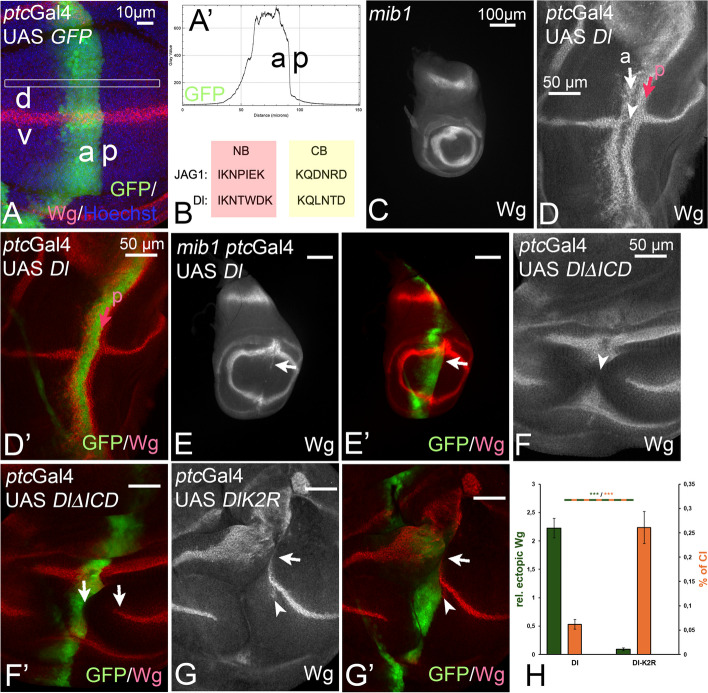
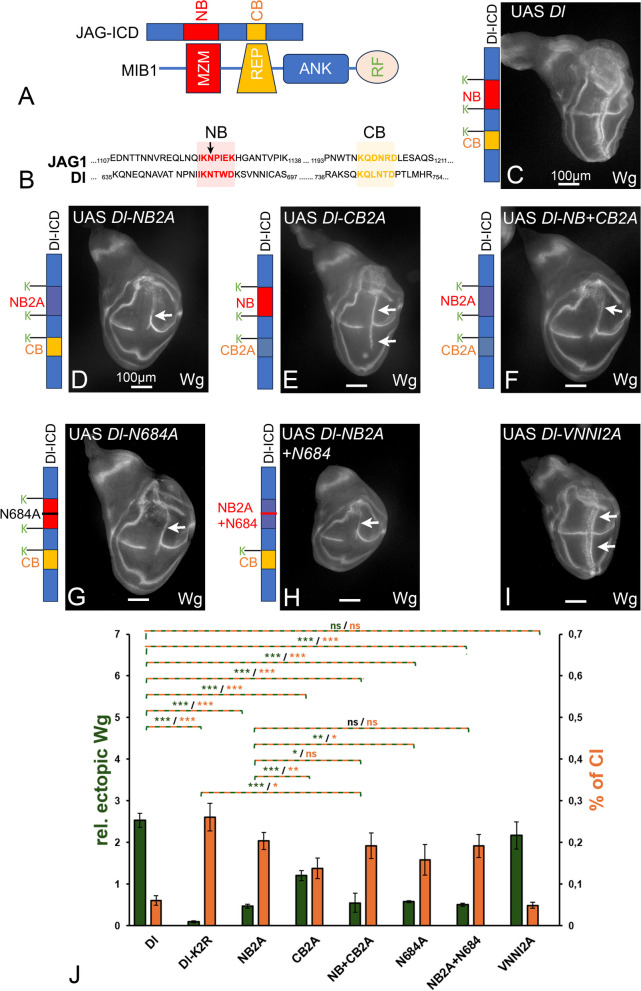
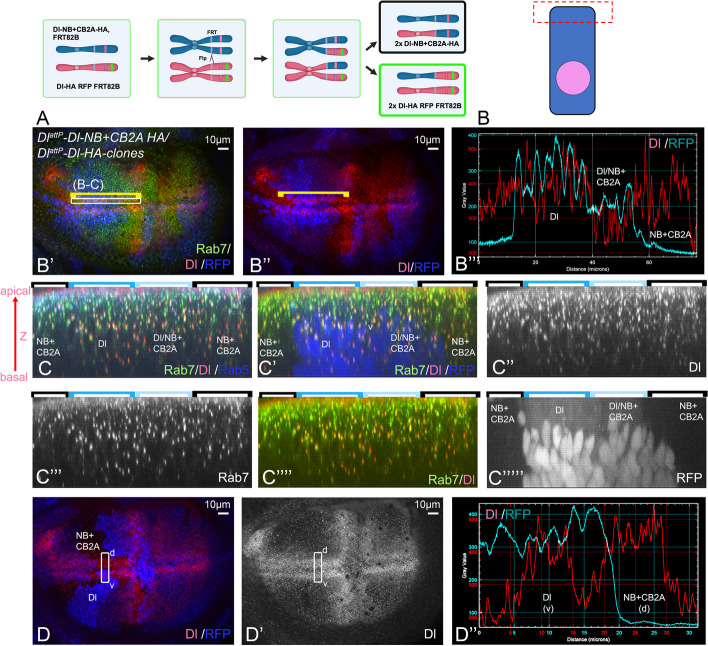
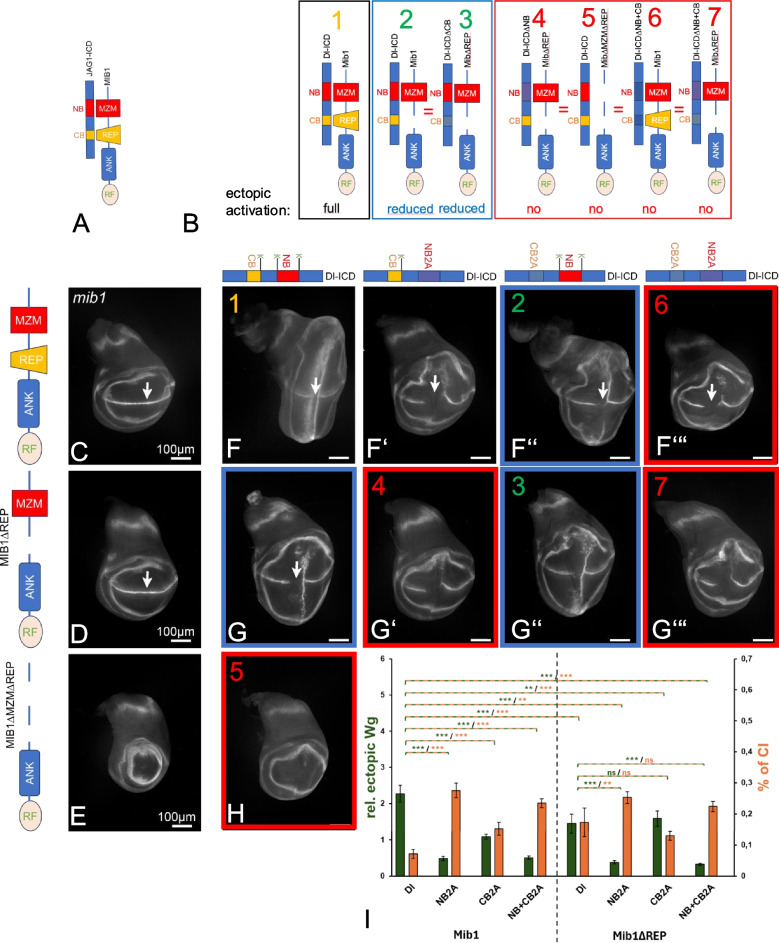
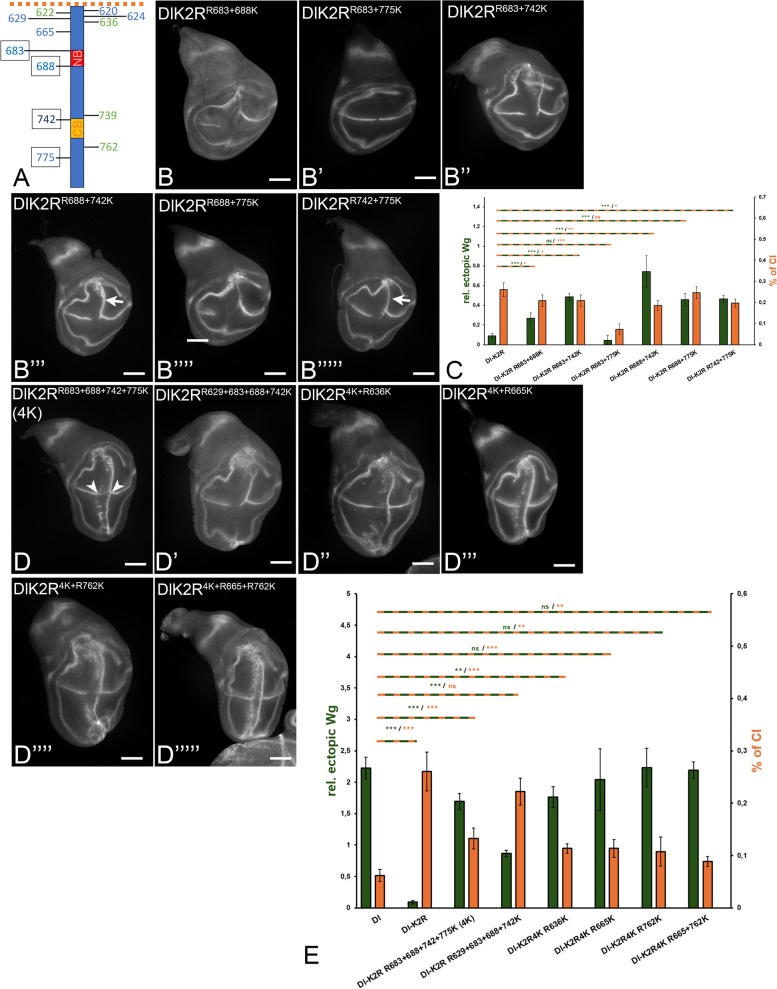
Background: Notch signalling plays a crucial role in many developmental, homoeostatic and pathological processes in metazoans. The pathway is activated by binding of the ligand to the Notch receptor, which changes the conformation of the receptor by exerting a pulling force. The pulling force is generated by the endocytosis of the interacting ligand into the signal-sending cell. Endocytosis of ligands requires the action of the E3 ligases Mindbomb1 (Mib1) and Neuralized (Neur) that ubiquitylate lysines (Ks) of their intracellular domains. It has been shown that human MIB1 binds JAGGED1 (JAG1) via a bipartite binding motif in its ICD. This interaction is required for the activation of JAG1. However, it is not known whether this bipartite binding mode is of general importance. It is also not rigorously tested whether it occurs in vivo. Moreover, it is not known whether Mib1 ubiquitylates specific Ks in the ICD of ligands, or is rather non-selective.
Results: We therefore investigated how Mib1 interacts with the Notch ligand Delta of Drosophila in an in vivo trans-activation assay and determined the Ks which are required for signalling. We show that the activation of Dl by Mib1 follows similar rules as has been found for mammalian MIB1 and JAG1. We present evidence that a combination of six Ks of the ICD is required for the full signalling activity of Dl by Mib1, with K742 being the most important one.
Conclusions: Altogether, our analysis further reveals the rules of Mib1-mediated DSL-ligand-dependent Notch-signalling.
Keywords: Cis-inhibition; DSL-ligands; Delta; Endocytosis; Mindbomb1; Notch-pathway; Ubiquitylation.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Not applicable. Consent for publication: All the authors of this manuscript consent to its publication as a research article in BMC Biology. Competing interests: The authors declare no competing interests.
Figures



References
-
- Siebel C, Lendahl U. Notch signaling in development, tissue homeostasis, and disease. Physiol Rev. 2017;97:1235–94. - PubMed
-
- Seib E, Klein T. The role of ligand endocytosis in notch signalling. Biol Cell. 2021. 10.1111/boc.202100009. - PubMed
-
- Schnute B, Troost T, Klein T. Endocytic trafficking of the Notch receptor. Adv Exp Med Biol. 2018;1066:99–122. 10.1007/978-3-319-89512-3_6. - PubMed
-
- Le Borgne R, Bardin A, Schweisguth F. The roles of receptor and ligand endocytosis in regulating Notch signaling. Development. 2005;132:1751–62. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
