

Hyperammonemia induces programmed liver cell death
- PMID: 40053595
- PMCID: PMC11887801
- DOI: 10.1126/sciadv.ado1648
Hyperammonemia induces programmed liver cell death
Abstract
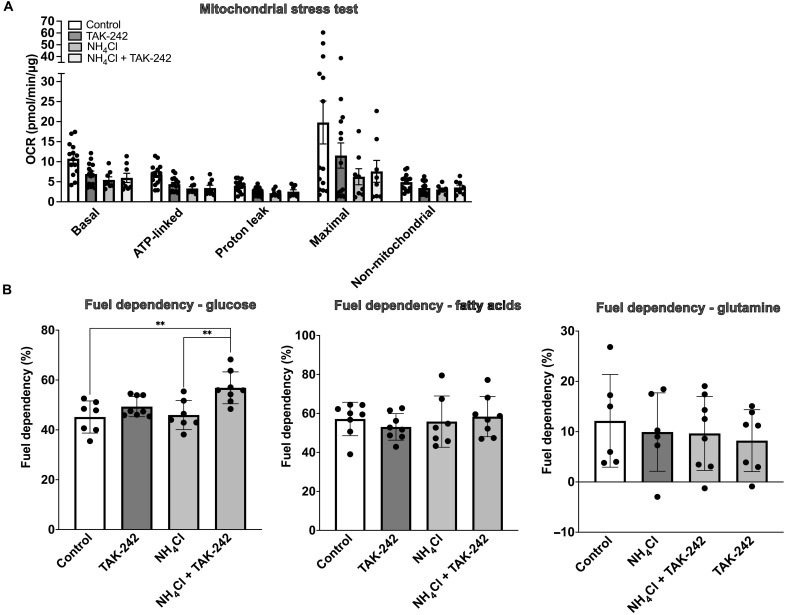
Hyperammonemia is common in liver cirrhosis and causally associated with hepatic encephalopathy development. Little is known about its hepatotoxic effects, which we aimed to characterize in this study. In a mouse model of chronic hyperammonemia without preexisting liver disease, we observed development of liver fibrogenesis and necroptotic cell death. Hyperammonemia also induced dysregulation of its main metabolic pathway, the urea cycle, as reflected by down-regulation of urea cycle enzyme protein expression and accumulation of its metabolites. Inhibition of receptor-interacting serine/threonine-protein kinase 1 (RIPK1) and its upstream inducer Toll-like receptor 4 (TLR4) protected against liver injury and further hyperammonemia. In clinically relevant rodent models of hyperammonemia (genetic ornithine transcarbamylase deficiency and bile duct ligation-induced cirrhosis), TLR4 inhibition reduced circulating ammonia. In conclusion, hyperammonemia induces liver fibrogenesis and RIPK1-mediated cell death, which is associated with urea cycle dysfunction. Inhibition of RIPK1 and TLR4 protects against hyperammonemia-induced liver injury and are potential therapeutic targets for hyperammonemia and chronic liver disease progression.
Figures






References
-
- Haussinger D., Regulation of hepatic ammonia metabolism: The intercellular glutamine cycle. Adv. Enzyme Regul. 25, 159–180 (1986). - PubMed
-
- Kaiser S., Gerok W., Haussinger D., Ammonia and glutamine metabolism in human liver slices: New aspects on the pathogenesis of hyperammonaemia in chronic liver disease. Eur. J. Clin. Invest. 18, 535–542 (1988). - PubMed
-
- Fabbri A., Marchesini G., Bianchi G., Bugianesi E., Bortoluzzi L., Zoli M., Pisi E., Unresponsiveness of hepatic nitrogen metabolism to glucagon infusion in patients with cirrhosis: Dependence on liver cell failure. Hepatology 18, 28–35 (1993). - PubMed
-
- Shangraw R. E., Jahoor F., Effect of liver disease and transplantation on urea synthesis in humans: Relationship to to acid-base status. Am. J. Physiol. 276, G1145-52 (1999). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
