

Mitochondrial damage in muscle specific PolG mutant mice activates the integrated stress response and disrupts the mitochondrial folate cycle
- PMID: 40057508
- PMCID: PMC11890779
- DOI: 10.1038/s41467-025-57299-3
Mitochondrial damage in muscle specific PolG mutant mice activates the integrated stress response and disrupts the mitochondrial folate cycle
Abstract
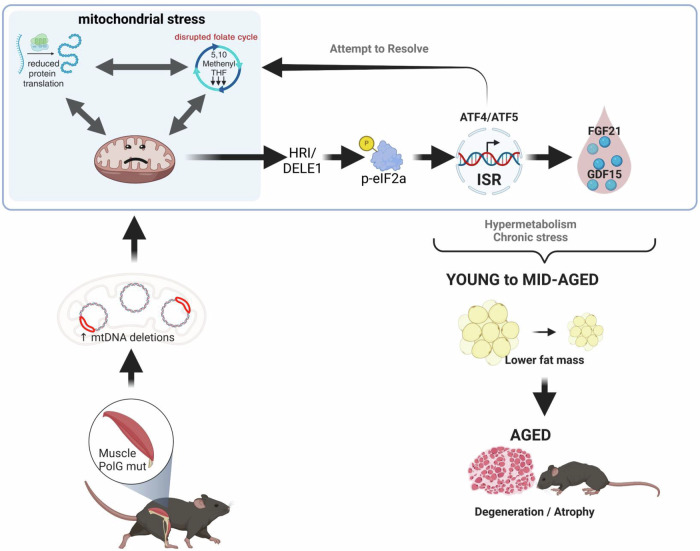
During mitochondrial damage, information is relayed between the mitochondria and nucleus to coordinate precise responses to preserve cellular health. One such pathway is the mitochondrial integrated stress response (mtISR), which is known to be activated by mitochondrial DNA (mtDNA) damage. However, the causal molecular signals responsible for activation of the mtISR remain mostly unknown. A gene often associated with mtDNA mutations/deletions is Polg1, which encodes the mitochondrial DNA Polymerase γ (PolG). Here, we describe an inducible, tissue specific model of PolG mutation, which in muscle specific animals leads to rapid development of mitochondrial dysfunction and muscular degeneration in male animals from ~5 months of age. Detailed molecular profiling demonstrated robust activation of the mtISR in muscles from these animals. This was accompanied by striking alterations to enzymes in the mitochondrial folate cycle that was likely driven by a specific depletion in the folate cycle metabolite 5,10 methenyl-THF, strongly implying imbalanced folate intermediates as a previously unrecognised pathology linking the mtISR and mitochondrial disease.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: BGD, STB and DCH together with the Baker Heart & Diabetes Institute declare they are pursuing protection of intellectual property (IP) around the use of proprietary folate cycle metabolite derivatives as therapeutics for mitochondrial diseases, and other degenerative conditions in which 1C-metabolism and the ISR are perturbed. All other authors declare no conflict of interest.
Figures




References
-
- Johns, D. R. Mitochondrial, D. N. A. and disease. N. Engl. J. Med.333, 638–644 (1995). - PubMed
-
- Falkenberg, M., Larsson, N. G. & Gustafsson, C. M. DNA replication and transcription in mammalian mitochondria. Annu. Rev. Biochem.76, 679–699 (2007). - PubMed
-
- Anderson, S. et al. Sequence and organization of the human mitochondrial genome. Nature290, 457–465 (1981). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
