

This is a preprint.
Comprehensive analysis of myeloid reporter mice
- PMID: 40060446
- PMCID: PMC11888320
- DOI: 10.1101/2025.02.24.639159
Comprehensive analysis of myeloid reporter mice
Abstract
Macrophages are a pivotal cell type within the synovial lining and sub-lining of the joint, playing a crucial role in maintaining homeostasis of synovium. Although fate-mapping techniques have been employed to differentiate synovial macrophages from other synovial myeloid cells, no comprehensive study has yet been conducted within the mouse synovial macrophage compartment. In this study, we present, for the first time, lineage tracing results from 18 myeloid-specific fate-mapping models in mouse peripheral blood (PB) and synovial tissue. The identification of synovial macrophages and monocyte-lineage cells through flow cytometry was further validated using cellular indexing of transcriptomes and epitopes by sequencing (CITE-seq) datasets. These findings provide a valuable methodological tool for researchers to select appropriate models for studying the function of synovial myeloid cells and serve as a reference for investigations in other tissue types.
Figures
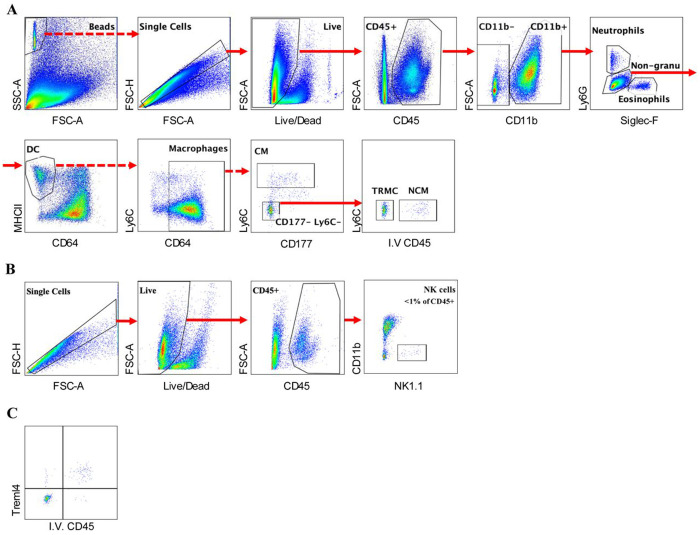

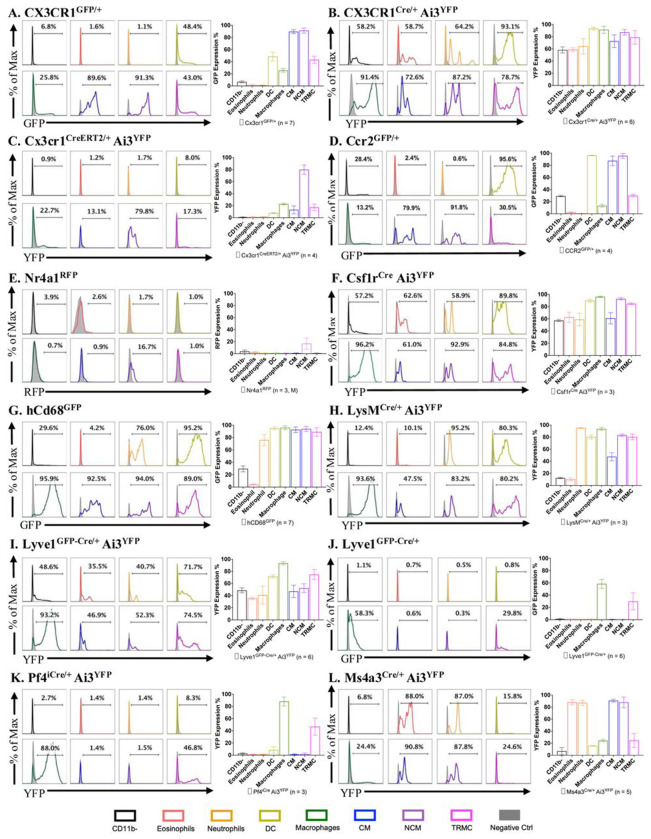




References
-
- Guilliams M., Mildner A. & Yona S. Developmental and Functional Heterogeneity of Monocytes. 49, 595–613 (2018). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources