

This is a preprint.
Distinct prelimbic cortex ensembles encode response execution and inhibition
- PMID: 40060503
- PMCID: PMC11888377
- DOI: 10.1101/2025.02.23.639736
Distinct prelimbic cortex ensembles encode response execution and inhibition
Update in
-
Distinct prelimbic cortex ensembles encode response execution and inhibition.Proc Natl Acad Sci U S A. 2025 Sep 16;122(37):e2505378122. doi: 10.1073/pnas.2505378122. Epub 2025 Sep 8. Proc Natl Acad Sci U S A. 2025. PMID: 40920924
Abstract
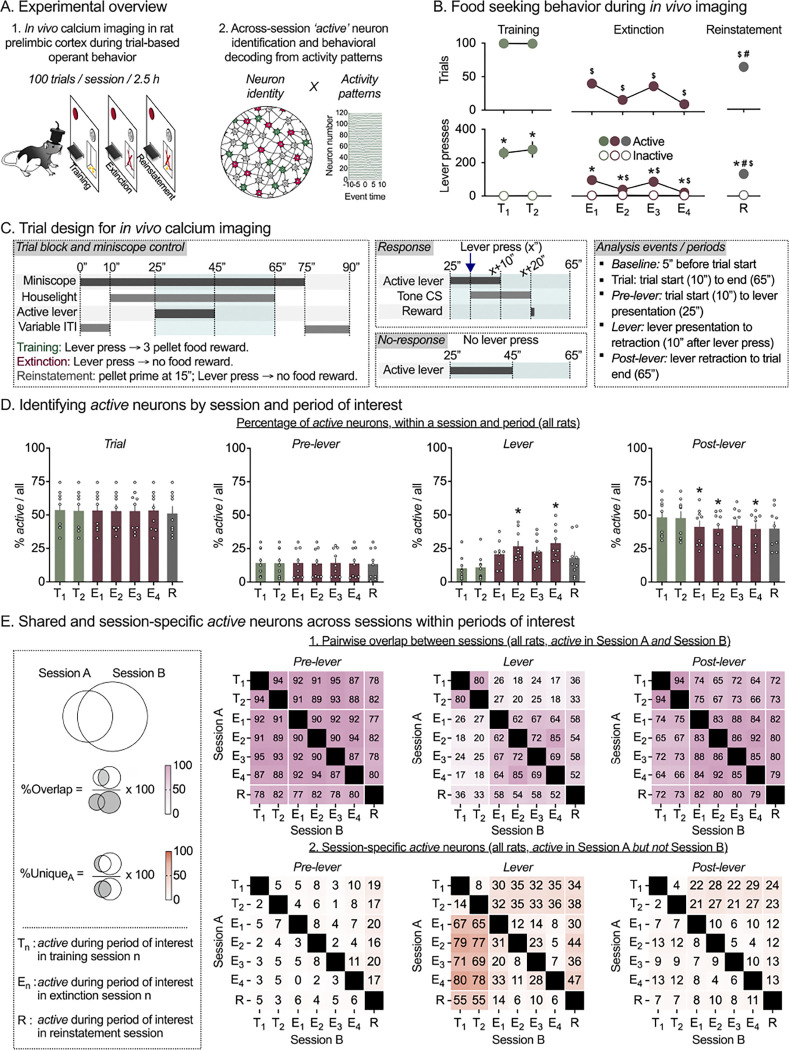
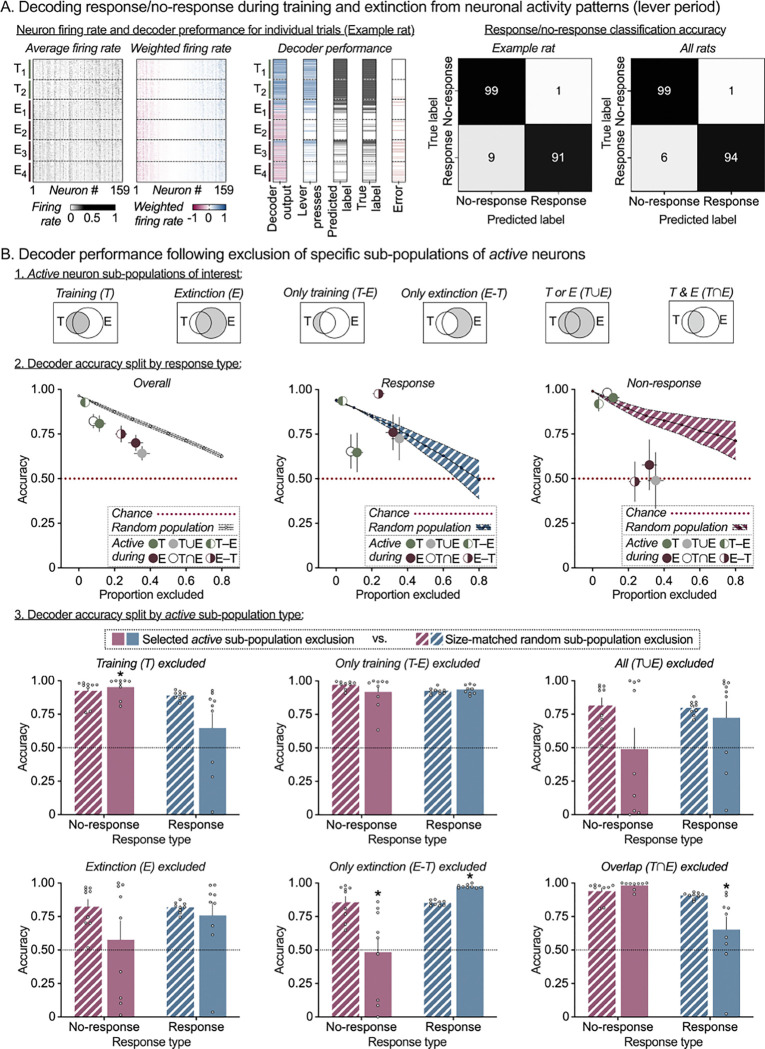
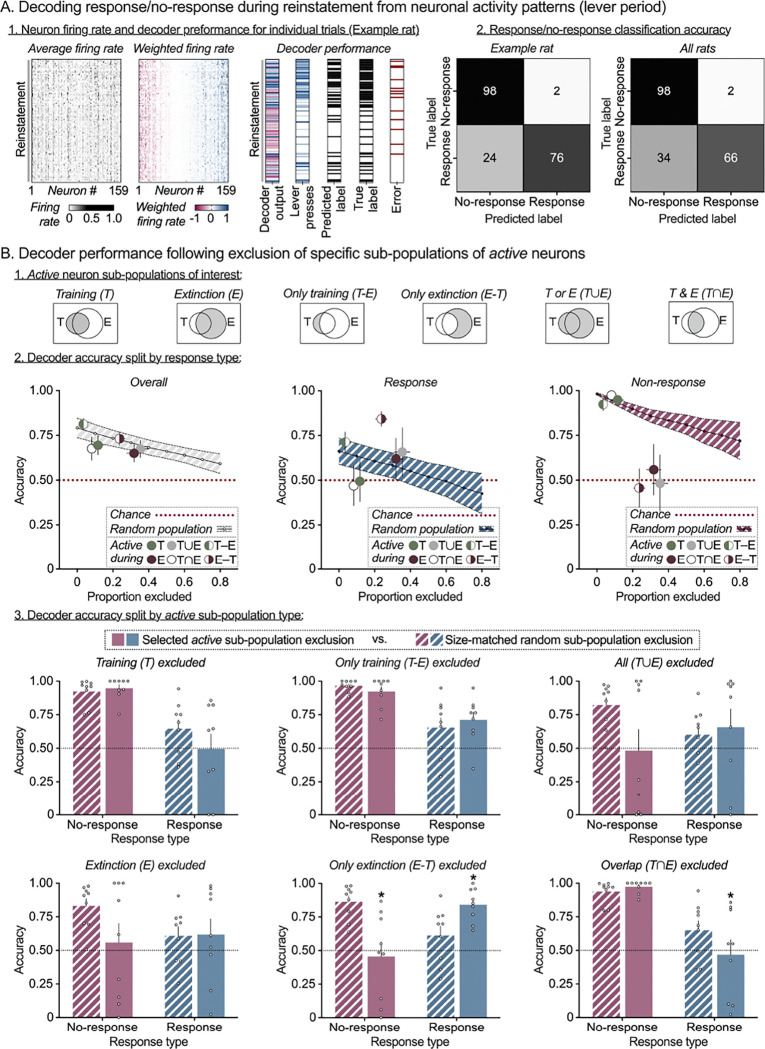
Learning when to initiate or withhold actions is essential for survival and requires integration of past experiences with new information to adapt to changing environments. While stable prelimbic cortex (PL) ensembles have been identified during reward learning, it remains unclear how they adapt when contingencies shift. Does the same ensemble adjust its activity to support behavioral suppression upon reward omission, or is a distinct ensemble recruited for this new learning? We used single-cell calcium imaging to longitudinally track PL neurons in rats across operant food reward Training, Extinction and Reinstatement, trained rat-specific decoders to predict trial-wise behavior, and implemented an in-silico deletion approach to characterize ensemble contributions to behavior. We show that operant training and extinction recruit distinct PL ensembles that encode response execution and inhibition, and that both ensembles are re-engaged and maintain their roles during Reinstatement. These findings highlight ensemble-based encoding of multiple learned associations within a region, with selective ensemble recruitment supporting behavioral flexibility under changing contingencies.
Conflict of interest statement
Competing Interests All authors declare that they do not have any conflicts of interest (financial or otherwise) related to the text of the paper.
Figures
References
-
- Dalley J. W., Cardinal R. N., Robbins T. W., Prefrontal executive and cognitive functions in rodents: neural and neurochemical substrates. Neurosci Biobehav Rev 28, 771–784 (2004). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous