

This is a preprint.
Distal mutations enhance catalysis in designed enzymes by facilitating substrate binding and product release
- PMID: 40060566
- PMCID: PMC11888230
- DOI: 10.1101/2025.02.21.639315
Distal mutations enhance catalysis in designed enzymes by facilitating substrate binding and product release
Update in
-
Distal mutations enhance catalysis in designed enzymes by facilitating substrate binding and product release.Nat Commun. 2025 Sep 30;16(1):8662. doi: 10.1038/s41467-025-63802-7. Nat Commun. 2025. PMID: 41027962 Free PMC article.
Abstract
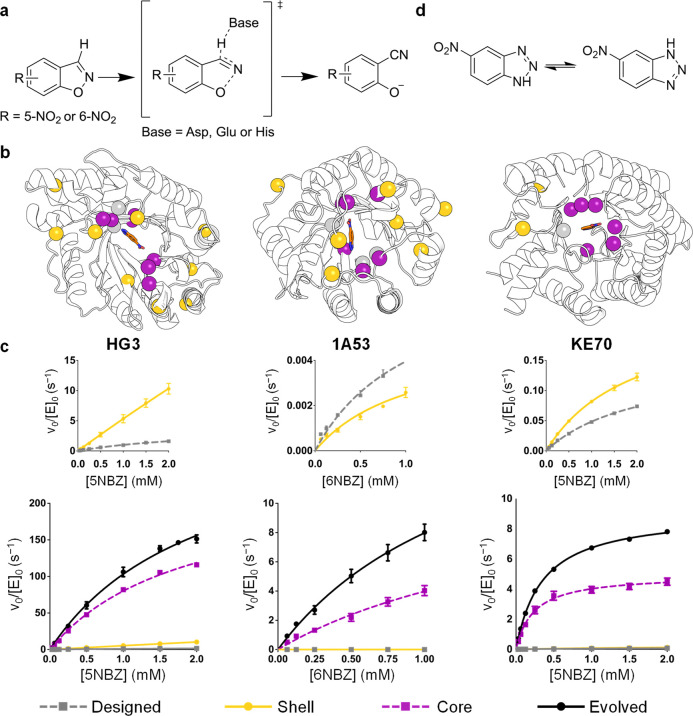
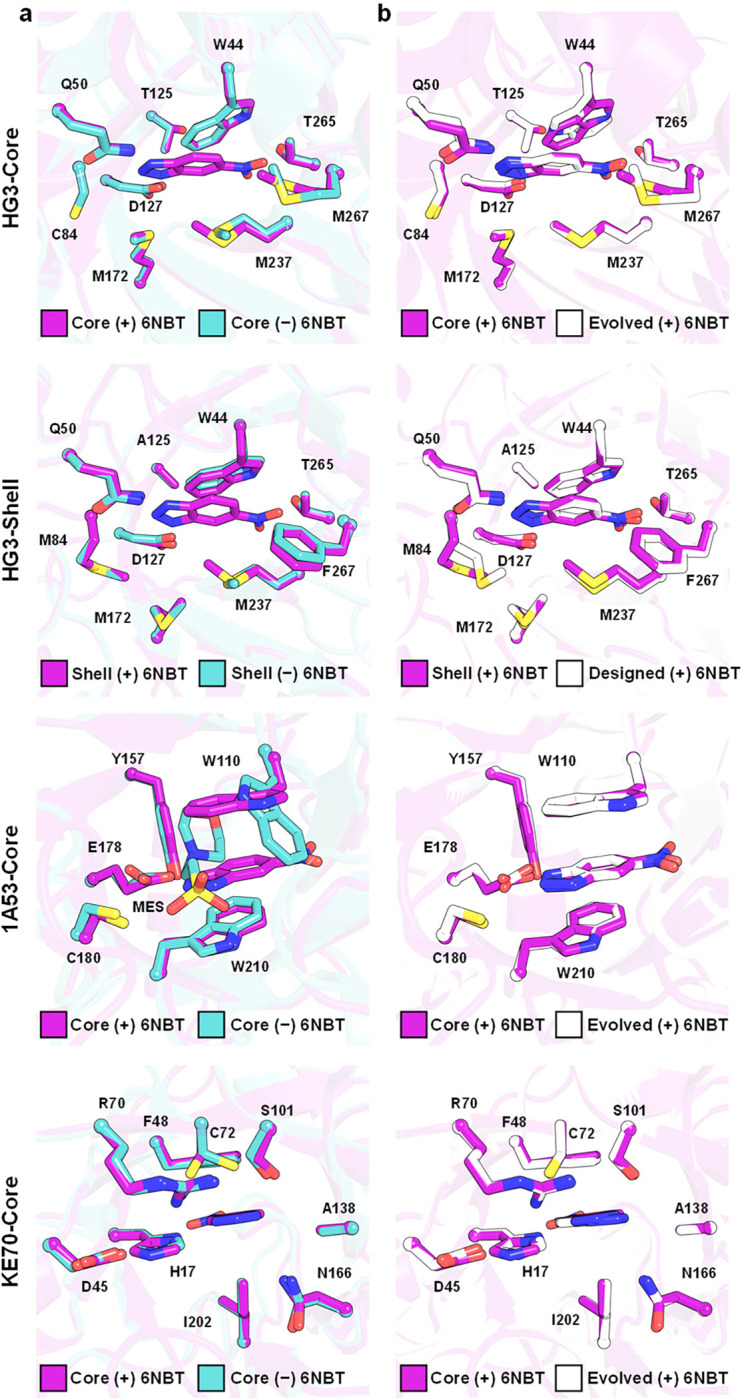
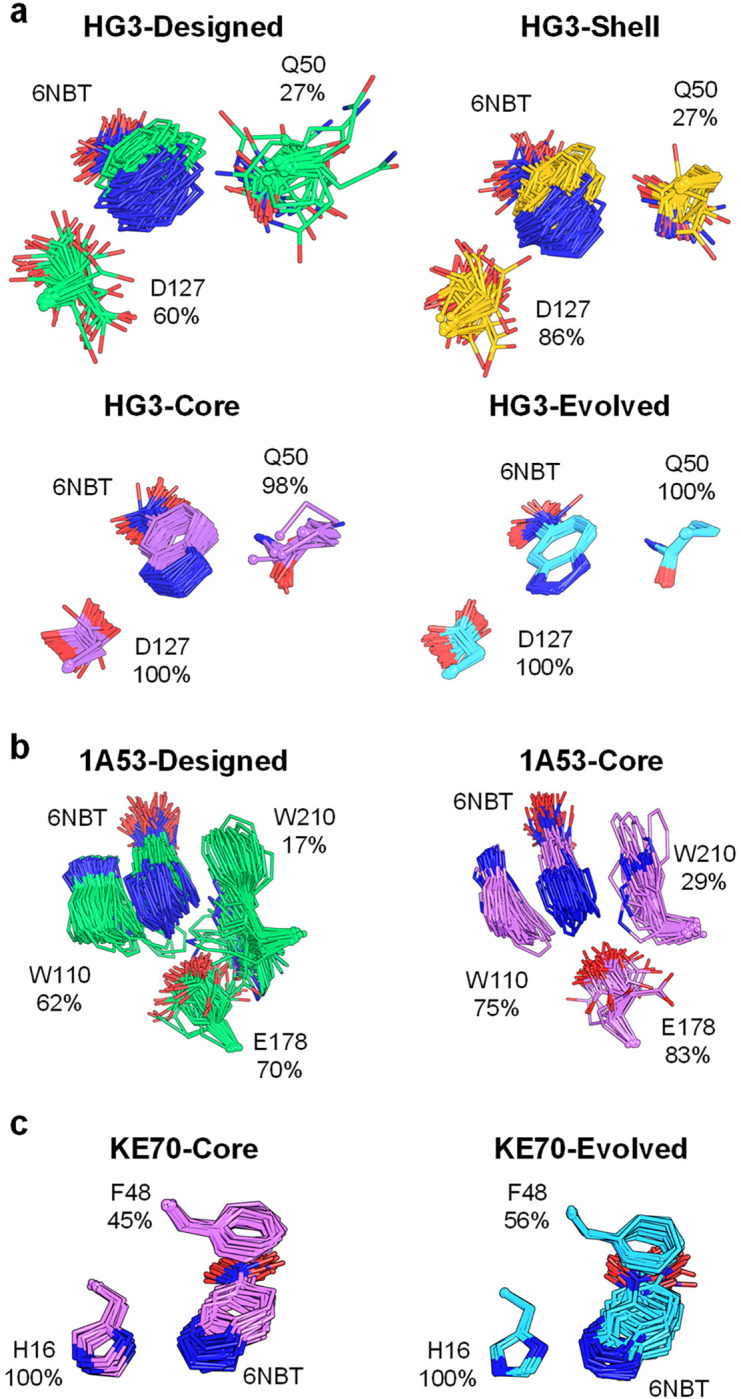
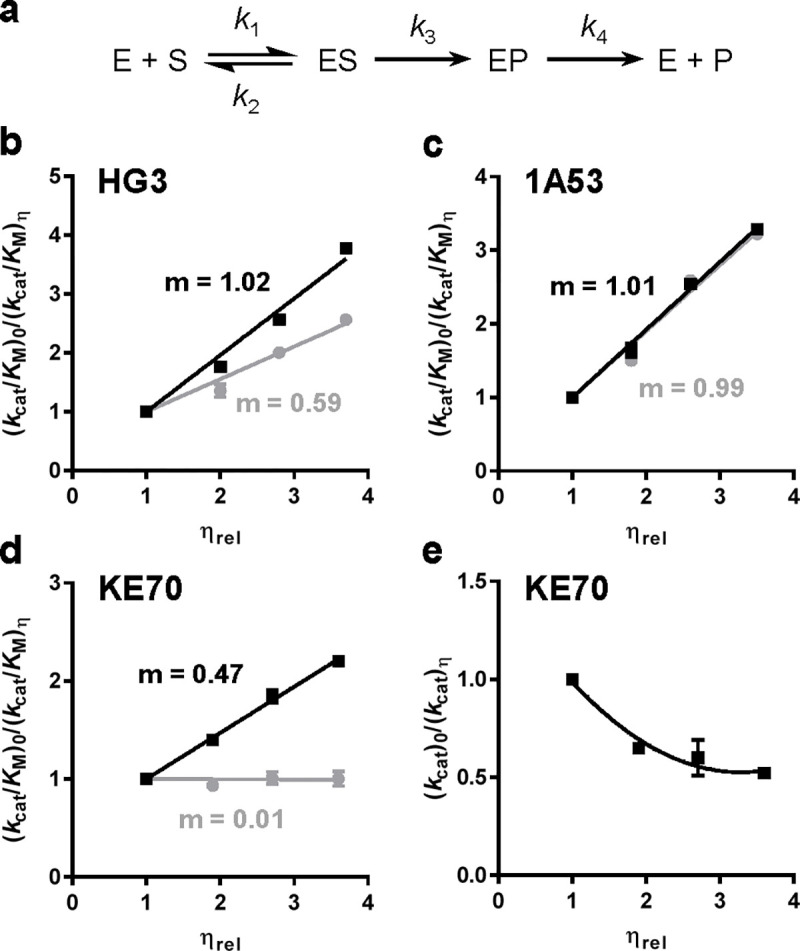
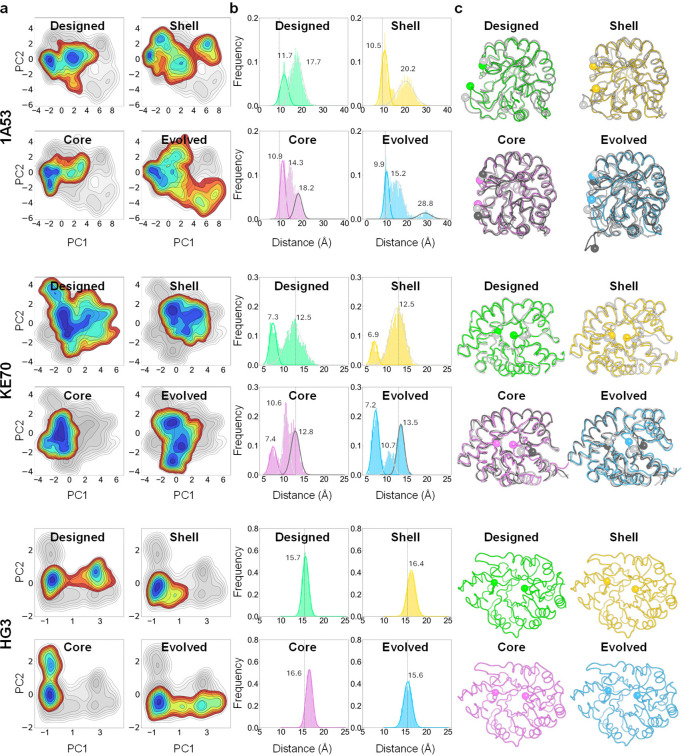
The role of amino-acid residues distant from an enzyme's active site in facilitating the complete catalytic cycle-including substrate binding, chemical transformation, and product release-remains poorly understood. Here, we investigate how distal mutations promote the catalytic cycle by engineering mutants of three de novo Kemp eliminases containing either active-site or distal mutations identified through directed evolution. Kinetic analyses, X-ray crystallography, and molecular dynamics simulations reveal that while active-site mutations create preorganized catalytic sites for efficient chemical transformation, distal mutations enhance catalysis by facilitating substrate binding and product release through tuning structural dynamics to widen the active-site entrance and reorganize surface loops. These distinct contributions work synergistically to improve overall activity, demonstrating that a well-organized active site, though necessary, is not sufficient for optimal catalysis. Our findings reveal critical roles that distal residues play in shaping the catalytic cycle to enhance efficiency, yielding valuable insights for enzyme design.
Keywords: Enzyme catalysis; X-ray crystallography; enzyme design; enzyme engineering; enzyme mechanisms; molecular dynamics.
Conflict of interest statement
Competing Interests The authors declare no competing interests.
Figures
References
-
- Du S. et al. Conformational ensembles reveal the origins of serine protease catalysis. Science 387, eado5068 (2025). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources